Anders als die Geophiliden nehmen Lithobiiden ihre Nahrung in Stücken auf, die mit Hilfe der bezahnten Mandibeln von der Beute abgerissen werden (MANTON 1965). Dadurch ist die Feststellung des Nahrungsspektrums über eine mikroskopische Analyse des Darminhalts möglich,
Für Darminhaltsanaiysen wurden nur Hundertfüßler verwendet, die frisch aus dem Freiland gefangen und in Alkohol oder Formalin 4 % konserviert waren. (Keine Tiere aus der Kempson-Extraktion). Der Darm der Tiere wurde frei präpariert, eröffnet, und der Inhalt auf einen Objektträger in Faures'sche Lösung übertragen. Die Herstellung der Faures'schen Lösung geschieht folgendermaßen:
5 cm³ Aqua dest., 30 g pulverisiertes Gummi arabicun, 20 ml Glycerin und 50 g Chloralhydrat werden unter Erwärmung vermischt. Anschließend wird die Lösung durch Glaswolle abfiltriert. (Angaben nach WEIDEMANN 1971)
Kleine Larvenstadien wurden auch nach Aufhellung in warmer Milchsäure als Ganzes eingebettet und zerquetscht. Das Durchmustern der Präparate geschah bei 160 x und 400 x Vergrößerung in Durchlicht und Phasenkontrast. Die gefundenen Strukturen wurden in einer Fotokartei zusammengestellt und mit Hilfe von Referenzpräparaten gequetschter Beutetiere (aus Kempson-Proben) verschiedenen Beutegruppen zugeordnet.
Tabelle 19 gibt die für bestimmte Taxa charakteristischen Strukturen an.
in Abb. 14 a - f ist dargestellt, in welchen Anteilen bestimmte Strukturen im Darm adulter Tiere der 6 häufigsten Lithobius-Arten im Gebiet FW zu finden waren. Alle Tiere wurden in der Streuschicht gefangen.
L. lusitanus valesiacus fällt durch hohen Prozentsatz von Tieren auf, deren Därme als leer eingestuft wurden (4 von 44 Individuen, Abb. 14 b). Tierische Fragmente wurden bei dieser Art nur in drei Individuen gefunden.
Die übrigen 5 Arten unterscheiden sich qualitativ fast nicht in ihrem Nahrungsspektrum. In der Häufigkeit, mit der bestimmte Beutegruppen angetroffen werden, gibt es anscheinend gewisse Unterschiede. Leider erlaubte die Seltenheit der meisten Arten nur kleine Stichprobengrößen ohne Beeinträchtigung des Untersuchungsgebietes. Deshalb ist nur der Unterschied im Nahrungsspektrum zwischen L. mutabilis und L. curtipes signifikant (P< 0,05, siehe Tab. 20).
Von einigen in der Streuschicht regelmäßig vorhandenen Tiergruppen wurden keine Überreste in den untersuchten Hundertfüßlern gefunden: Es handelt sich hierbei um Staphyliniden, Elateridenlarven, onychiuride Collembolen und Fliegenlarven. Es ist möglich, dass Einzelindividuen aus diesen Gruppen in die Kategorie "andere Tiere" eingestuft wurden, jedoch auch dann ist ihre Repräsentanz im Nahrungsspektrum der Lithobiiden wesentlich geringer, als nach der Häufigkeit im Waldboden zu erwarten wäre. Das gilt auch für die Milben, die in der Kempsonextraktion erheblich zahlreicher als die entomobryiden Collembolen sind, im Darminhalt der Lithobiiden aber seltener als Collembolen vorkommen.
Vergleicht man das Nahrungsspektrum der Lithobiiden im Winter (Dez.-März) mit der übrigen Zeit des Jahres (Abb. 15), so zeigt sich für 2 Kategorien ein Unterschied in der Antreffhäufigkeit, der mit großer Wahrscheinlichkeit nicht zufällig ist (Tab. 21 a- c): Im Winter werden vermehrt Pilzhypen und -sporen aufgenommen, während Pilzmücken hauptsächlich im Frühjahr, Sommer und Herbst gefressen werden, Tatsächlich wurden im Darm auch überwiegend Reste von Pilzmücken Imagines gefunden. (Im Winter kommen Mycetophiliden hauptsächlich als Larven vor.
Erstaunlich ist, dass der Prozentsatz von Tieren mit leerem Verdauungstrakt im Jahreszeitvergleich gleich bleibt. Es bleibt unklar, ob dies durch eine unverminderte Fressaktivität im Winter, besonders lange Verweildauer der Nahrung im Darm bei Kälte oder (wie JOSSE & TESTERINK (1977) bei Collembolen festgestellt haben) durch eine Periode des Nahrungsmangels im Sommer zu erklären ist.
Da Dipteren keine wesentliche Rolle für die Unterschiede im Nahrungsspektrum der Arten spielen, halte ich es für berechtigt, für die folgende Nischenbetrachtung die Gesamtproben heranzuziehen.
(pi = Anteil der Nahrungskategorie i an der Summe aller Nahrungsnachweise für die Art.)
Diese einfache Formel wurde gewählt, da die Datenbasis sowieso nur grobe Vergleiche zulässt.
Die verschiedenen Arten scheinen eine sehr ähnliche Nutzungsbreite der Nahrungsgruppen zu haben. Betrachtet man nur die tierische Beute, besteht der größte Unterschied zwischen L. curtipes und L. mutabilis (letzte Spalte Tab. 22), er ist aber geringer als die jahreszeitliche Differenz der Nahrungs-Nischenbreite für die Summe aller adulten Lithobiiden. (Der hohe NB-Wert von L. dentatus basiert auf einer zu kleinen Datenmenge, um differenzierte Vergleiche zuzulassen.)
Juvenile Stadien von L. mutabilis und L. curtipes haben ein engeres Nahrungsspektrum als die adulten Tiere (Abb. 16, Tab. 23). Die Unterschiede in der Nischenbreite sind größer als zwischen den Adulten der verschiedenen Arten und übertreffen auch die jahreszeitliche Differenz der Nischenbreite (Tab. 22).
Der Vollständigkeit halber wurden die Berechnungen nicht nur für die 7 Gruppen von Beutetieren sondern auch für alle 40 Darminhaltskategorien durchgeführt. Ich will mich in den folgenden Betrachtungen auf die für die Beutetiergruppen berechneten Werte beschränken, da unklar ist, ob die anderen Bestandteile des Darminhalts gezielt als Nahrung aufgenommen werden (LEWIS 1965).
Tabelle 24 zeigt die spezielle Nischenüberlappung zwischen den Adulten der 6 Lithobius-Arten (nach MÜHLENBERG 1976):
(pij = Anteil der Nahrungskategorie i an der Nahrung der Art j; pihAnteil der Nahrungskategorie i an der Nahrung der Art h.)
Die Überlappung ist groß (der Sonderfall L. lusitanus wurde bereits oben erwähnt). Die durchschnittliche Überlappung der Nahrungsrische einer Art mit der aller anderen ist sehr ähnlich (Tab. 24, äußerste rechte Spalte und unterste Zeile). Die Arten L. macilentus und L. muticus weisen zwar etwas tiefere Werte auf als die 3 anderen, es ist aber leicht möglich, dass diese Unterschiede auf Zufall beruhen.
Wesentlich geringere NU-Werte ergeben sich beim paarweisen Vergleich der verschiedenen Altersklassen von L. mutabilis und L. curtipes (Tab. 25). Bemerkenswert ist die geringe Nischenüberlappung zwischen adulten L. curtipes und den größengleichen Stadien PL 3-5 von L. mutabilis. Ein Blick auf Abb. 16 zeigt, dass dies auf die größere Häufigkeit von Milben in der Nahrung dieser Stadien von L. mutabilis zurückzuführen ist, Die Wahrscheinlichkeit für ein zufälliges Auftreten dieses Häufigkeitsunterschieds ist kleiner als 5 % (X²-Test).
Die Berechnung der durchschnittlichen Nischenüberlappung zeigt noch deutlicher, dass sich die Altersklassen von L. mutabilis in der Nahrungsnutzung ferner stehen als die verschiedenen Arten untereinander. (Tab. 26)
HURLBERT (1978) und PETRAITIS (1979) weisen darauf hin, dass bei der Berechnung von Nischenparametern die Häufigkeit (besser noch: Verfügbarkeit) der Ressourcen zu berücksichtigen sei und schlagen entsprechende Formeln vor.
Probeweise berechnete ich die Nischenbreite für L. mutabilis, L. curtipes und L. mutabilis PL 3-5 nach dem von PETRAITIS angegebenen Verfahren. Das Häufigkeitsverhältnis der Beutegruppen ermittelte ich nach den Ergebnissen der Kempson-Extraktion (Tab. 6, Spalte: "ohne Handauslese"). Wie Tabelle 27 zeigt, ändern sich die Ergebnisse durch den Petraitis-Index nicht; die Verhältnisse werden sogar eher noch etwas deutlicher. MAIORANA (1978 b) zeigte in einer Untersuchung der Nahrung von Salamandern, dass es darüber hinaus nicht immer berechtigt ist, die Häufigkeit einer Tiergruppe in der Extraktion mit ihrer Verfügbarkeit für den Räuber gleichzusetzen. In der vorliegenden Untersuchung stammen alle Tiere aus dem gleichen Gebiet. Man kann also grob annehmen, dass die mittlere Verfügbarkeit der Beute für alle Arten bzw. Stadien gleich ist. Da die geringen Stichprobengrößen sowieso nur grobe Vergleiche zulassen, scheint mir in diesen Fall die Anwendung der einfachen Nischenindices von COLWELL & FUTUYMA (1971) berechtigt zu sein.
Eine den Verhältnissen bei L. mutabilis ähnliche Entwicklung des Nahrungsspektrums im Verlauf der Individualentwicklung zeigt Lithobius variegatus LEACH in einem Buchen-Eichenwald in Südengland (ROBERTS 1957). Auch bei dieser Art besteht das Beutespektrum in den frühen Entwicklungsstadien hauptsächlich aus Collembolen, um sich dann später vor allem um Enchytraeen, Spinnen und Milben zu erweitern. Die aus den Daten von ROBERTS (1957: Tab. 37) berechnete durchschnittliche intraspezifische Nischenüberlappung von L. variegatus (Stadienzusammenfassung wie bei L. mutabilis) entspricht mit NT = 0,38 dem für L, mutabilis berechneten Wert.
Bei so wenig selektiven Räubern, wie Lithobiiden es zu sein scheinen, stellt sich die Frage, ob Nahrungsanalysen aus einem Gebiet oder Habitat für eine Population in anderer Umgebung etwas aussagen können. Zur Beantwortung dieser Frage verglich ich die Ergebnisse der Studien von ROBERTS (1957) und LEWIS (1965) an Lithobius variegatus und L. forficatus untereinander sowie meine Ergebnisse zu L. muticus mit denen von ROBERTS (1957).
[Anmerkung: Möglicherweise handelt es sich um verschiedene Arten. Dafür spricht auch die von ROBERTS für "seinen" L. muticus angegebene geringe Trockentoleranz (Überlebenszeit 9,8 Stunden bei 66 % rH), die sich nicht mit der Feststellung verträgt, dass L. muticus allgemein eher trockenere Habitate als L. mutabilis bevorzugt (ALBERT 1982, LOKSA 1966, Kap. 4 dieser Arbeit)."]
ALBERT (im Druck a) führte mit Lithobius mutabilis und L. curtipes Laborversuche zur Beutewahl durch. Sie beobachtete, dass L. mutabilis auch Beutetiere annahm, die für L. curtipes zu groß waren, stellte aber sonst keinen Unterschied in der Beutewahl zwischen den beiden Arten fest.
Eine Reihe der laut ALBERT gern gefressenen Beutegruppen konnte ich nicht im Darm der von mir untersuchten Lithobiiden entdecken (v, a. Onychiurus spec., Diplura, Aphidae), andererseits fanden sich nicht selten Überreste von Hornmilben, die in den von ALBERT durchgeführten Versuchen nur ungern und in der Regel nur nach einer Hungerperiode genommen wurden. Der Vergleich der Beutewahlversuche mit dem aus dem Freiland festgestellten Nahrungsspektrum unterstützt die oben ausgesprochene Vermutung, dass das Vorkommen einer Tiergruppe in der Kempson-Extraktion nicht mit ihrer Verfügbarkeit für die Lithobiiden gleichgesetzt werden darf.
1.Der große Anteil leerer Därme bei L. lusitanus valesiacus bestätigt die in Kapitel 2 getroffene Feststellung, dass diese Art ihren Lebensraum nicht in der freien Streuschicht hat.
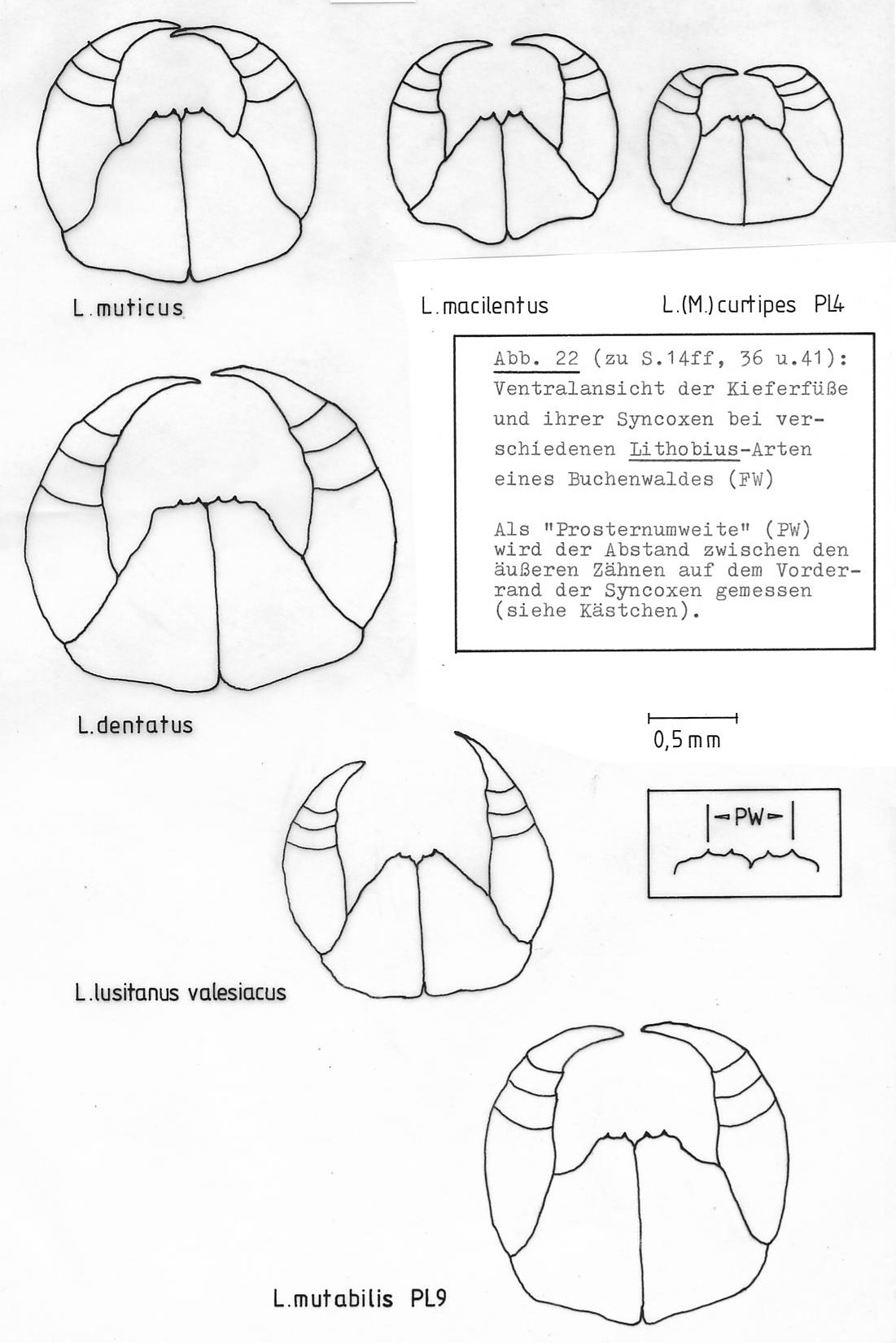
2.Für die drei häufigsten Chilopodenarten der Streuschicht 1äßt sich eine ökologische Sonderung in Bezug auf die Nahrung annehmen: Strigamia acuminata gehört als Geophilide einem anderen Ernährungstypus an, der weiche, fleischige Nahrung bevorzugt (VERHOEFF 1925, MANTON 1965). Zwischen L. mutabilis und L. (M.) curtipes besteht ein deutlicher Größenunterschied, besonders in Bezug auf die für die Ernährung wichtige Bezahnung des Coxosternuns der Kieferfüße (siehe auch Abb. 22). Größengleiche Entwicklungsstadien der beiden Arten haben eine geringere Überlappung ihrer Nahrungsspektren als die Adulten, deren Prosternumweite sich um den Faktor 1,3 unterscheidet. Laborbeobachtungen legen nahe, dass die Arten sich auch im Beutefangverhalten unterscheiden: L. mutabilis nimmt häufig schon ab Stadium PL1 dargebotene immobilisierte Drosophila-Fliegen von der Pinzette, während auch ausgewachsene L. (M.) curtipes regelmäßig mit Flucht reagieren und die Beute erst verzehren, wenn sie einige Zeit gelegen hat.
3.Für die übrigen Chilopodenarten der Streuschicht (L. muticus, L. dentatus und mit Einschränkung L. macilentus) konnte die Analyse des Darminhalts keine Erklärung für ihre Koexistenz in der Chilopoden-Community erbringen.
4.Die Tatsache, dass die intraspezifische Überlappung der Nahrungsnischen von L. mutabilis-Altersklassen geringer ist, als die interspezifische der 6 untersuchten Hundertfüßler, lässt die Vermutung aufkommen, dass die intraspezifische Konkurrenzvermeidung (oder besser - verminderung) eine wesentliche Rolle bei dieser im Gebiet sehr häufigen Art spielt. In der Schlussdiskussion werde ich auf diesen Gedanken im Zusammenhang mit der Phänologie und Altersstruktur der Population zurückkommen.
Die vorhandenen Daten lassen keine Entscheidung über die Frage zu, ob die festgestellten Unterschiede im Nahrungsspektrum sekundärer Ausdruck anderer Sonderungsmechanismen (z. B. Habitatwahl) sind, oder ob sie als unmittelbare Strategie zur Konkurrenzverminderung verstanden werden können. (S. dazu MAIORANA 1978).
Nahrungsspektrum der Lithobiiden
(1) Alle Lithobius-Arten der Streuschicht haben ein weites Beutespektrum, das sich vor allem aus Collembolen, Enchyträen, Milben, kleinen Spinnen und Dipteren zusammensetzt.
(2) Alle Arten scheinen Detritus und Pilzhyphen und -sporen aufzunehmen.
(3) Verschiedene Altersklassen des dominanten Lithobius mutabilis stehen sich im Nahrungsspektrum ferner als die verschiedenen Arten der Streuschicht untereinander.
(4) Juvenile Stadien haben ein engeres Beutespektrum als adulte Tiere.
(5) Lithobiiden fressen sich gegenseitig.
(6) Ergebnisse von Darminhaltsanalysen an Lithobiiden dürfen nur mit großer Vorsicht auf andere Gebiete übertragen werden,
Hinweis: Tabellen und Abbildungen können mit [Mausklick rechts -> Link in neuem Fenster öffnen] so aufgerufen werden, dass sie parallel zum Text geöffnet bleiben
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}