Zur Feststellung eindeutiger Kausalbeziehungen zwischen den Chilopoden und ihrer Umwelt ist die kontrollierte Veränderung vermuteter Einflussgrößen im Experiment unerlässlich. Unter sonst natürlichen und kontrollierten Bedingungen sollten Parameter wie das Nahrungsangebot, der Räuberdruck, die Populationsdichte etc. variiert werden, um ihre Auswirkungen auf das Zusammenleben der Chilopoden studieren und nachweisen zu können. Synökologische Experimente mit Hundertfüßlern unter natürlichen oder halbnatürlichen Bedingungen werfen jedoch einige Probleme auf:
- Chilopoden lassen sich nicht direkt beobachten.
- Chilopoden lassen sich nicht mit Fallen repräsentativ erfassen.
- Die allein erfolgreiche Auslese der Chilopoden aus ihrem Substrat führt immer zur Zerstörung des jeweiligen Habitats. Dadurch sind wiederholte Untersuchungen der selben Fläche nicht möglich. Eine zu große Zahl von Stichproben führt zur Gefahr des "Aufbrauchens" der Versuchsfläche. Dasselbe Problem gilt auch für eine Kontrolle des Nahrungsangebots der Chilopoden.
Damit entfällt die Möglichkeit einer störungsfreien Versuchsüberwachung, und Zeitreihen lassen sich nur durch Versuchswiederholungen mit verschiedener Laufzeit gewinnen.
Den daraus erwachsenden Anforderungen konnte ich im Rahmen der vorliegenden Untersuchung nicht gerecht werden. Die hier dargestellten Versuche und Ergebnisse sind deshalb auch nur als orientierende Vorversuche aufzufassen, die einen ersten Eindruck von den methodischen Problemen und von der Art der zu erwartenden Resultate geben können.
Wie reagiert die Chilopoden-Artengemeinschaft auf die Verknappung von Ressourcen? Wie wirkt sich eine Veränderung des Ressourcenangebots auf das Arten- und Dominanzspektrum der Lithobiiden aus?
Diesen Fragen versuchte ich in Labor- und Freilandversuchen mit Hundertfüßlern nachzugehen.
In 20x20 x 7 cm³ Polystyrol-Klarsichtboxen wurde die Entwicklung künstlich zusammengesetzter Lithobius-Populationen verfolgt. Die Schachteln wurden mit einer ca. 5 mm dicken Schicht aus feuchtem Sand und einer ca. 50 mm dicken Schicht aus hitzeextrahiertem und wiederbefeuchteten Streumaterial gefüllt. Die künstliche Streuschicht bestand in ihren unteren 10 - 45 mm aus feinerem Material (Siebdurchgang bei 2 cm Maschenweite), darüber aus totem Laub. Zur Nahrungsversorgung wurde einmal vor Einsetzen der Hundertfüßler die Laubstreu in den Schachteln mit frischem, sorgfältig auf Chilopoden kontrollierten, Waldstreu-Gesiebe beimpft (Maschenweite des Siebes 3 mm). Es wurden nur adulte und subadulte Lithobiiden eingesetzt. Die Kunststoffschachteln waren mit Deckeln versehen, die in ihrer Mitte ein Loch von 15 mm Ø zur Lüftung hatten. Außer gelegentlichem Ausgleich des durch Wägen ermittelten Feuchtigkeitsverlustes fand keine Beeinflussung der Populationen bis zur Extraktion im Kempson-Apparat statt.
Die Ergebnisse der Versuche sind in Tabelle 42 zusammengefaßt. Es lassen sich folgende Feststellungen machen:
(1) Unabhängig von der Ausgangsdichte herrscht in den gemischten Populationen am Schluss der Experinente eine Dichte von ca. 5 Tieren pro Schachtel. Auf den Quadratmeter umgerechnet liegt diese Dichte (120/m²) im Bereich der auch im Freiland gefundenen Abundanz (115/m²; siehe Legende zu Tab. 42).
(2) Die Dichte der "Beute"-Gruppe (Collembolen, Milben, Enchyträen, Dipteren, Kleinspinnen) entwickelt sich bis zu Werten, die nur wenig unter den Freiland-Dichten liegen. In den Laborschachteln stellt sich ungefähr das gleiche Verhältnis "Beute"/Lith. ein wie im Freiland. (Nimmt man nur epimorphe und adulte Lithobiiden als Bezug, beträgt das Beuteangebot im Labor die Hälfte des im Freiland ermittelten Wertes.) Kontrollschachteln ohne Lithobiiden hatten am Versuchsende höhere Beutedichten als solche mit Hundertfüßlern (nicht signifikant; t-Test, P = 0,07). Die Lithobiiden-Enddichte ist signifikant mit der "Beute"-Zahl in der jeweiligen Schachtel korreliert (r = 0,77, P< 0,01).
(3) Für L. mutabilis scheinen die Bedingungen in den Schachteln ungünstiger zu sein als für L. (M.) curtipes. (Größere Verlustanteile bei reinen Populationen.)
(4) L. mutabilis scheint der interspezifischen Konkurrenz in den Laborschachteln weniger gewachsen zu sein als L. (M.) curtipes (und weitere Arten?). (In gemischten Populationen wesentlich höhere Verluste bei L. mutabilis als bei den anderen Arten.)
Während der Versuche zeigte sich, wie schwierig es ist, Freilandbedingungen im Labor zu simulieren. Das wurde z. B. am Problem der Streubefeuchtung deutlich. Im Freiland wird der Feuchtegradient im Boden immer wieder durch Niederschläge oder Tau aufgehoben oder verringert. Die Laborschachteln waren dagegen diesen Schwankungen nicht ausgesetzt, was dazu führte, dass bei Versuchsende ein Teil der Schachteln am Boden sehr feucht, in den oberen Blattschichten jedoch ausgesprochen trocken war. Auch das Nahrungsangebot in den Schachteln ließ sich - abgesehen vom ersten Beimpfen mit Mesofauna - nicht kontrollieren.
Junge Larven als Anzeichen für eine Fortpflanzung der Chilopoden unter den Laborbedingungen fand ich nur in den im Oktober 1979 angesetzten Versuchen. Auf ein eingesetztes Weibchen entfielen dabei im Schnitt 1,3 Larven. Die Vermehrungsrate war also unnatürlich klein. Außerdem ist nicht auszuschließen, dass die Larven aus Eiern geschlüpft waren, die mit dem Impfmaterial eingeschleppt wurden. Es bleibt also festzuhalten, dass die Laborpopulationen sich nicht oder kaum fortpflanzten.
Aufgrund dieser ungelösten Probleme, wie auch der kleinen Zahl von Versuchswiederholungen sind die Experimente nur als Vorversuche aufzufassen, die den Rahmen der technischen Probleme abstecken, deren Ergebnisse aber lediglich Vermutungen zulassen und weiter überprüft werden müssen.
Zu diesen Ergebnissen zählt die sich andeutende Regulation der Schachtel-Besatzdichte durch das Beuteangebot sowie der Befund, dass die dominante Art L. mutabilis unter verstärkten Konkurrenzbedingungen möglicherweise der sympatrischen Art L. (M.) curtipes unterlegen ist. Bei einer weiteren Überprüfung dieser Frage müsste vor allem abgesichert werden, dass nicht die gegenüber dem Freiland veränderten Klima- und Habitatverhältnisse in den Laborschachteln die Ursache für die hohe Mortalität von L. mutabilis sind.
Im Rahmen eines anderen Forschungsprojekts (MÜHLENBERG et al. 1984) waren in den Untersuchungsgebieten FWL und. ZAL mehrere Boden-Photoeklektoren installiert. Mit den ca. 7 - 9 cm tief in den Boden eingegrabenen Eklektoren wird eine Bodenfläche von 1 m² gegen die Umgebung abgegrenzt (siehe Abb. 32). Tiere im Eklektor können sich in der im Boden eingegrabenen Barberfalle und im Fangaufsatz auf der Eklektorspitze fangen. Die hauptsächlich für produktionsbiologische Untersuchungen eingesetzten Eklektoren (FUNKE 1971) lassen sich auch als Freilandversuche interpretieren. Folgende Faktoren sind unter den Bodeneklektoren verändert:
(1) Die Populationen sind gegen die Außenwelt abgegrenzt.
(2) Niederschläge werden durch das Zeltdach des Eklektors weitgehend abgehalten.
(3) Die Bodenfläche unter dem Eklektor ist von Laubfall ausgeschlossen.
(4) Es ist anzunehmen, dass Spinnen und Carabiden durch die Fallen im Eklektor weggefangen werden und damit ihr Einfluß unter dem Eklektor reduziert wird.
Ich untersuchte die Lithobiiden-Fauna unter je zwei Eklektoren der Gebiete FWL und ZAL. Die Eklektoren waren am 14.03.1979 installiert worden, die Untersuchungen erfolgten zusammen mit dem Abbau der Fanggeräte in der dritten Maiwoche 1980. Ich verglich die Laubstreu innerhalb und außerhalb des Eklektors durch Streudickenmessung mit einer Stricknadel sowie durch Messung der Streukompressibilität mit einem Standardgewicht. Zwei Streuproben von 25 x 25 cm² innerhalb und zwei Proben außerhalb des Eklektors wurden im Kempson-Apparat extrahiert. Die übrige Laubstreu unter dem Eklektor wurde im Labor sorgfältig gesiebt und Lithobiiden sowie potentielle Lithobiidenräuber (große Spinnen und Staphyliniden) von Hand ausgelesen.
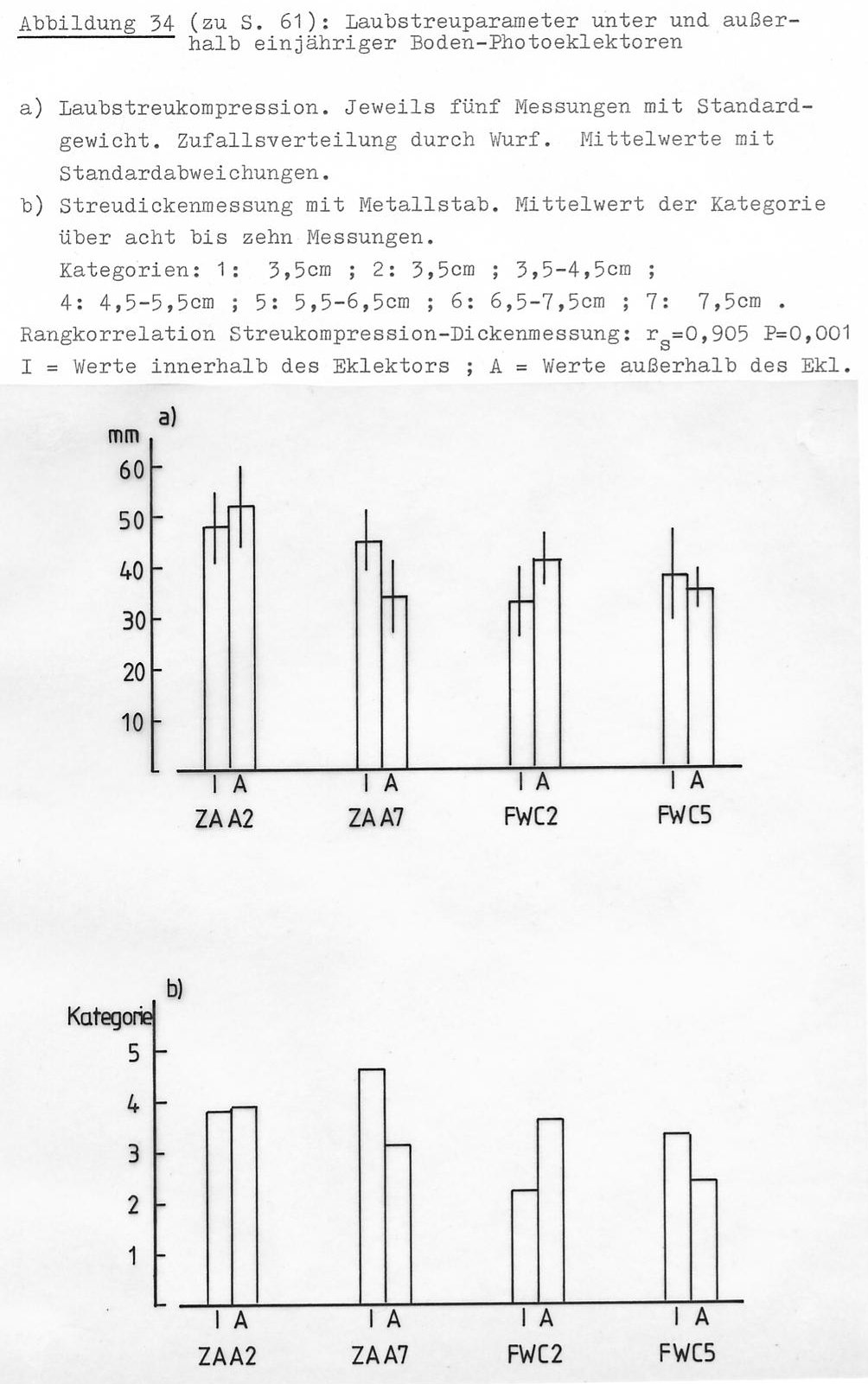
Die Laubstreu unter den Bodeneklektoren unterscheidet sich optisch von der der Umgebung (Abb. 33):
- Die Blätter haben innen einen mehr graubraunen Farbton; außen sind sie eher rötlichbraun.
- Außerhalb der Eklektoren ist die Laubstreu stärker aufgelockert als innerhalb. (Die Blätter liegen außerhalb mehr horizontal-vertikal gemischt.)
- Zum Zeitpunkt der Untersuchung (Mai) enthielt die Bodenauflage des Waldes infolge des Laubtriebes viele Blattknospenschuppen, die unterhalb des Eklektors natürlich nicht zu finden waren. Trotzdem zeigte sich in den Messwerten von Streudicke und -kompression kein Unterschied zwischen innen und außen (Abb. 34). Ich hatte den Eindruck, dass der Boden unter den Eklektoren trockener war als außerhalb. Messungen wurden hierzu leider nicht durchgeführt.
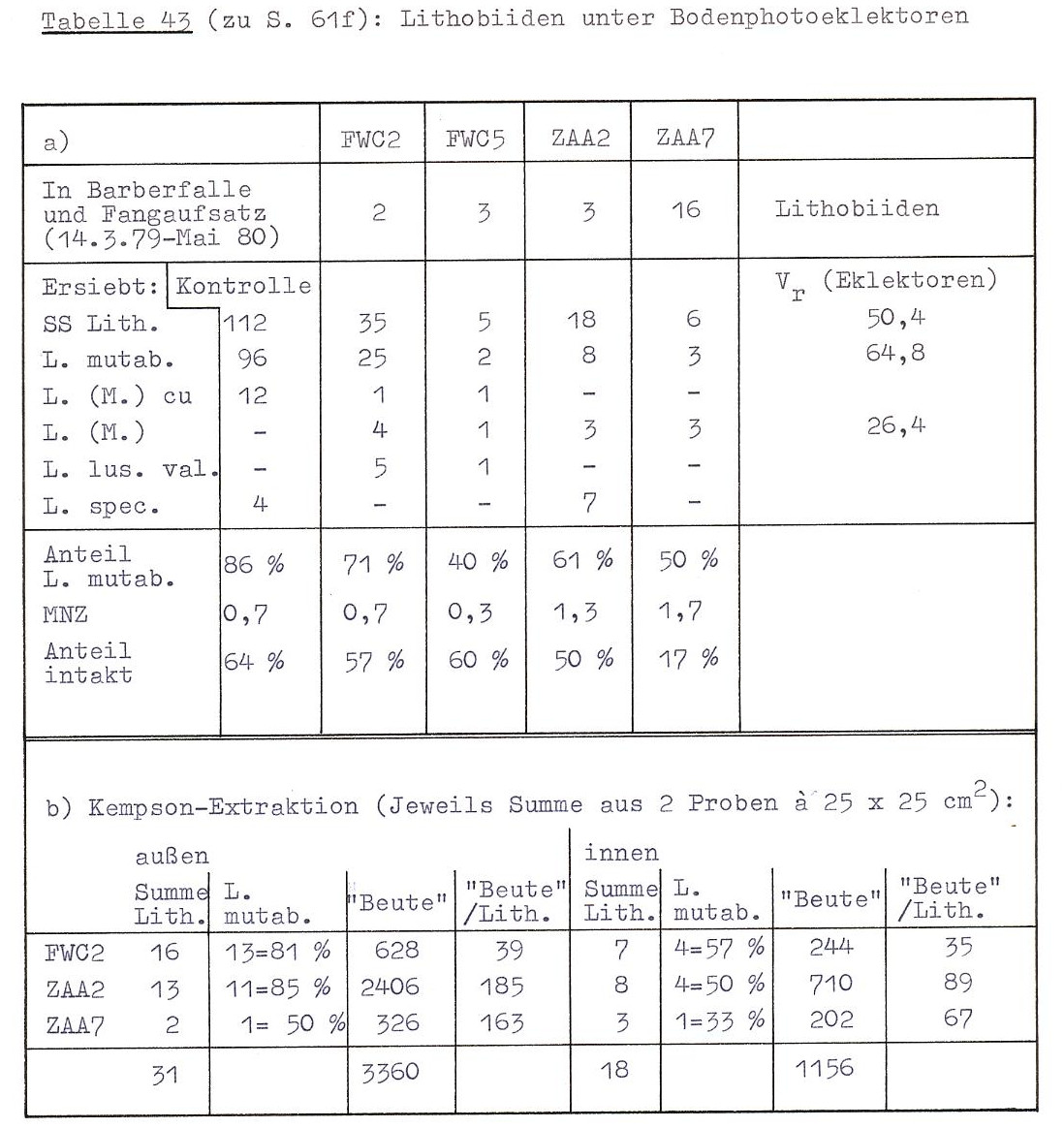
Tabelle 43 fasst einige Informationen über die Lithobiidenpopulationen unter den Eklektoren zusammen. Folgende Ergebnisse sollen besonders herausgehoben werden:
(1) Nach einem Jahr ist die Lithobiidendichte unter den Eklektoren deutlich verringert. Das zeigt sich sowohl beim Vergleich der ausgesiebten Streu mit Siebproben aus normalen Flächen des Gebietes FW (Tab. 43 a), wie auch beim Innen- Außenvergleich der Kempsonproben (Tab. 43 b). Dieses Ergebnis ist nicht durch die Existenz der Eklektorfallen zu erklären, wie aus einem Blick auf die Fangzahlen deutlich wird (1. Zeile Tab. 43 a). Die besonderen Verhältnisse des Eklektors ZAA7 erklären sich wahrscheinlich dadurch, dass im und um den Eklektor eine besonders hohe Spitzmaus-Aktivität herrschte. Dieser Eklektor war auch durch Grabgänge mit der Umgebung verbunden.
(2) Bodenproben aus dem Eklektor enthielten weniger "Beute" (= Collembolen, Milben, Enchyträen, Dipteren, Kleinspinnen) als die jeweils außerhalb der Eklektoren genommenen Proben. Ebenso ist die Anzahl "Beute" pro Lithobius im Eklektor geringer als außerhalb (Tab. 43 b).
(3) Unter den Eklektoren ist die Dominanz von Lithobius mutabilis geringer als außerhalb. Dies wird deutlich an einem signifikant niedrigeren Anteil von L. mutabilis an der Lithobius-Gesamtzahl im Eklektor. (Tab. 43 b; t-Test, paarweise Beobachtungen: t = 4,84, P< 0,05.)
(4) Im Gebiet FWL taucht unter den Eklektoren die sonst dort in der Streuschicht extrem seltene Art L. (M.) crassipes auf. Gleichzeitig geht der Anteil von L. (M.) curtipes an der Lithobiidenpopulation zurück.
Ganz allgemein machen diese Ergebnisse deutlich, dass nach einem Jahr die Lebensbedingungen unter einem Bodeneklektor verändert sind. Die geringe Dichte der Mesofauna spricht dafür, dass eine Ressourcenverknappung eingetreten ist, was auch wegen des fehlenden Laubfalls nicht anders zu erwarten wäre. Die Tatsache, dass die Streudicke sich dennoch nicht von der außerhalb der Eklektoren unterscheidet spricht gegen ein einfaches "Aufbrauchen" des Detritus als Ursache der Ressourcenverknappung. Möglicherweise sind die fehlenden Niederschläge für eine allgemeine Verschlechterung der Lebensbedingungen und damit auch eine geringere Stoffumsatzrate verantwortlich.
Es lässt sich nicht entscheiden, ob der Rückgang der Chilopodenzahl unter den Eklektoren eine Folge der verringerten Beutezahl ist, oder ob er durch die gleichen Faktoren verursacht wird, die auch für die Abnahme der Mesofauna verantwortlich sind. Das Auftauchen von Lithobius (Monotarsobius) crassipes und der Rückgang von L. (M.) curtipes in den Eklektoren des Gebietes FWL ließe sich als eine Folgeerscheinung größerer Trockenheit unter dem Eklektor interpretieren. Nach JEEKEL 1963, 1977 ist L. curtipes in den Niederlanden auf Moore und Flussufer beschränkt während L. crassipes mehr trockene Wälder bevorzugt. Im Steigerwald fand ich L. crassipes auch auf Wiesen.
Die nachlassende Dominanz von L. mutabilis unter den Eklektoren stützt die aufgrund der Laborversuche ausgesprochene Vermutung einer relativ geringen Konkurrenzkraft dieser Art. (L. mutabilis hat eine größere Trockenresistenz als L. curtipes - siehe 8.25 -, deshalb scheint mir größere Trockenheit unter den Eklektoren in diesen Fall keine ausreichende Erklärung zu sein.)
Im März/April 1979 wurden 12 Flächen (1 m²) durch einen 7 bis 8 cm tief in den Boden eingegrabenen 25 cm hohen Ring aus Hart-PVC gegenüber der Umgebung isoliert. Die Flächen wurden ursprünglich mit der Absicht eingerichtet, in ihnen experimentell die Dichte der Lithobiiden und ihrer Räuber zu manipulieren. Dieses Vorhaben wurde aber wegen zu großer methodischer Schwierigkeiten wieder fallengelassen. Möglich erschien jedoch eine Untersuchung der folgenden Fragen:
- Wirkt die beschriebene Eingrenzungsmethode isolierend auf die Lithobiiden?
- Welche Wirkung hat ein Eingrenzen von Teilpopulationen auf ihre Entwicklung?
- Wie wirkt sich eine Veränderung der Laubstreu auf die Chilopodenfauna aus?
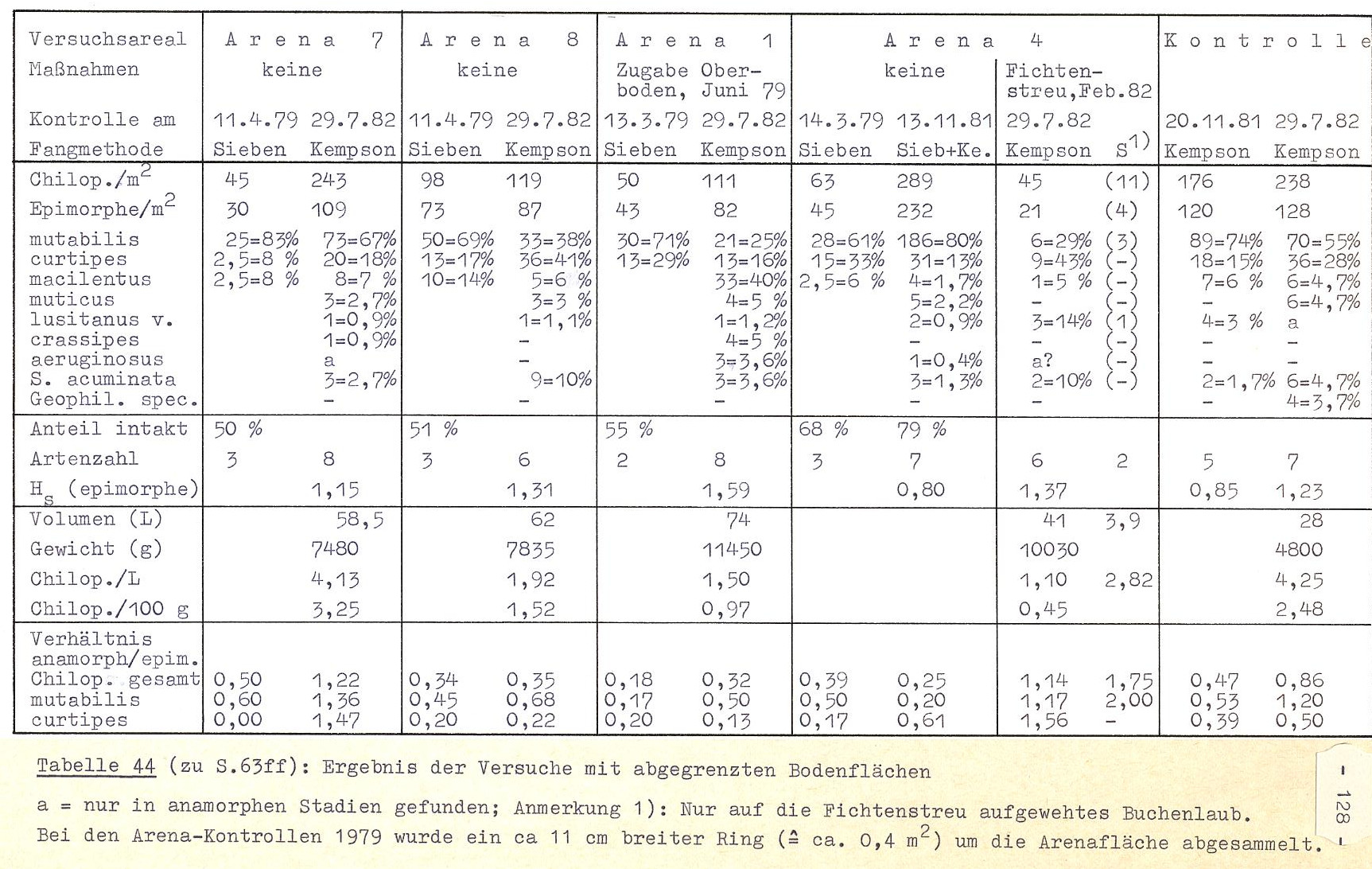
Fünf Arenen kamen zur Auswertung. Ein Überblick über die durchgeführten Behandlungen ist in Tabelle 44 gegeben. Ohne weitere Veränderungen außer dem Eingraben des Begrenzungsringes wurde die Bodenstreu einer Fläche (Arena 4) nach 2 1/2 Jahren (im November 1981) von Hand und anschließend durch Hitzeextraktion ausgelesen. Im Juli 1982 wurde die Streu zweier weiterer seit 3 1/2 Jahren nur horizontal abgegrenzter, sonst aber unbehandelter Versuchsflächen ohne Handauslese im Kempson-Apparat extrahiert.
Im Juni 1979 wurde zu einer Fläche (Arena 1) der hitzeextrahierte Oberboden (L-, F- und obere Humusschicht) von 1/2 m² Waldfläche aus dem Gebiet zugefügt. Im Juli 1982 (nach Verlauf von 3 Jahren) wurde der Oberboden der Arena 1 im Kempson-Apparat extrahiert.
In die Arena 4 (deren Boden seit November 1981 ohne Streuauflage war) gab ich Mitte Februar 1982 die hitzeextrahierte Bodenstreu von einem Quadratmeter Fichtenwaldboden des Gebietes FE. Die Kontrolle der Besiedlung erfolgte im Juli 1982 (nach Ablauf von 5 1/2 Monaten) ebenfalls durch Hitzeextraktion des Oberbodens im Kempson-Apparat. Auf die Fichtenstreu an einer Seite der Arena aufgewehte Buchenblätter sammelte ich getrennt und las zwischen ihnen enthaltene Chilopoden von Hand aus (Spalte S in Tabelle 44).
Als Kontrolle sammelte ich 1979 vor dem Eingraben der PVC-Ringe in einem Ring von 11 cm Breite außen um die Arena die Laubstreu, aus der ich die Lithobiiden im Labor von Hand auslas. Außerdem wurden parallel zu den Erhebungen der Jahre 1981 und 1982 je 8 Kempson-Bodenproben à 1/16 m² aus der Umgebung der Arenen genommen.
Die Versuchsergebnisse sind in Tabelle 44 zusammengefasst.
a) Unveränderte Flächen.
Die Versuchsflächen, in denen außer dem Eingrenzen keine weiteren Maßnahmen vorgenommen wurden, bieten ein sehr uneinheitliches Bild. Die 1981 abgesammelte Arena 4 zeichnete sich durch eine extrem hohe Siedlungsdichte der Lithobiiden aus. Die Werte liegen deutlich oberhalb des 95 % Vertrauensbereiches der Kontrolle von 1981 (95 % VB für Gesamtchilopoden = 86-222 Ind/m², für Epimorphe = 54-186 Ind/m²). Besonders Lithobius mutabilis war mit großer Dominanz vertreten. Die Narbenhäufigkeit war mit 79 % intakten Tieren besonders gering. Dabei war die Dichte chilopodenfressender Spinnen mit 30/m² normal. In den beiden 1982 abgesammelten Arenen 7 und 8 bestätigte sich dieses Ergebnis nicht. Gegenüber der Kontrolle 1982 ist die Lithobiidendichte in den Versuchsflächen eher verringert. Die Werte liegen in einem Fall (Arena 7) innerhalb des 95%-Vertrauensbereichs der Kontrolle, im anderen (Arena 8) tiefer. (95 % VB der Kontrolle 1982 für Gesamtchilopoden = 172-304 Ind/m², für Epimorphe = 92-164 Ind/m².) Auch die Ergebnisse der Kontrollzählungen 1979 (Absammeln eines Ringes um die Arenafläche vorm Eingraben) zeigen keine Beziehungen zu den Fangergebnissen in den jeweiligen Arenen nach 2 bzw. 3 Jahren. Ebenso ist das Zahlenverhältnis zwischen anamorphen und epimorphen Lithobiiden in den verschiedenen Flächen sehr unterschiedlich, lässt aber keinen Zusammenhang mit anderen Parametern erkennen.
Es ist sehr schwer, diese widersprüchlichen Resultate zu interpretieren. Möglicherweise drückt sich in ihnen ein gewisses Eigenleben der mehr oder weniger isolierten Teilpopulationen aus. Auf jeden Fall verdeutlichen diese Flächen die Schwierigkeiten, auf die die Durchführung von Freilandexperimenten im Waldboden stößt, denn als Kontrollflächen zu einem Experiment mit z. B. Manipulation der Räuberdichte böten sie ein äußerst unbefriedigendes Bild.
b) Streuvermehrung
Auf den Boden der Arena 1 wurde im Juni 1979 nicht nur Laub sondern auch Erde, Humus und halbzersetzte organische Substanz gegeben. Deshalb fand sich beim Absammeln eine besonders starke Schicht lockeren Bodens mit viel Humus und in Zersetzung befindlichem Material unter der normal entwickelten Laubschicht. Wie Tabelle 44zeigt, zeichnete sich diese Versuchsfläche 3 Jahre nach der Streuvermehrung durch eine besonders hohe Diversität aus. Es ist die einzige Fläche, in der Lithobius macilentus die häufigste Art ist. Bemerkenswert ist auch das gemeinsame Vorkommen aller drei Arten der Untergattung Monotarsobius (curtipes, crassipes, aeruginosus).
Es muss offen bleiben, ob durch die Veränderung des Bodens in Arena 1 tatsächlich mehr Nischen für die Lithobiiden geschaffen wurden, oder ob das Ergebnis nur die Momentaufnahme des Übergangs von einer Artengemeinschaft zu einer anderen ist, die Diversitätserhöhung also störungsbedingt ist (siehe z. B. LEBRUN 1976).
c) Veränderung der Laubstreu: Fichten- statt Buchenstreu
Während des Winters 1981/82 war der Boden der Arena 4 ohne Streubedeckung dem Frost ausgesetzt. Es ist deshalb unwahrscheinlich, dass beim Absammeln im November 1981 entkommene Lithobiiden dort genügend Schutz zum Überwintern fanden. Dennoch wurden im Juli 1982, 5 1/2 Monate nach Zufügen der Fichtenstreu, 56 Chilopoden von dieser Fläche gefangen. Es hat also offensichtlich trotz der Flächenbegrenzung eine Neubesiedlung der Bodenstreu stattgefunden. Die Tiere können über Regenwurm- oder Mausgänge, mit verwehten Blättern und abgefallenen Zweigen in die Fläche gelangt sein.
Die bei Versuchsende festgestellte Dichte der epimorphen Lithobiiden stimmt gut mit dem Wert überein, der am Ursprungsort der Fichtenstreu im selben Substrat festgestellt wurde (Gebiet FE: 26 Chilopoden gesamt/m², 20 Epimorphe/m².) Die wesentlich höhere Zahl der anamorphen Larven in der Versuchsarena zeigt, dass die eingewanderten Tiere sich erfolgreich fortgepflanzt hatten.
Die Dominanz von L. (M.) curtipes in der Arena entspricht ebenfalls den ursprünglich in der Streu im Gebiet FE vorgefundenen Verhältnissen. Die Dominanz ist jedoch im Fichtenwald mit 74 % wesentlich stärker ausgeprägt. L. mutabilis ist unter den Versuchsbedingungen stärker präsent als ursprünglich im Fichtenwald.
Vergleicht man das Vorkommen von L. mutabilis in den aufgewehten Buchenblättern (Spalte S der Tabelle 44) mit dem in der Fichtenstreu der Versuchsfläche, so zeigt sich allerdings eine eindeutige Bevorzugung des Buchenlaubes: In der Fichtenstreu fanden sich 0,32 Individuen pro Liter, im Buchenlaub sind es 1,54 pro Liter. Lithobius mutabilis ist also zwischen den Buchenblättern ungefähr 5 mal häufiger als in der Fichtenstreu. (Die angegebenen Zahlen beziehen sich auf alle Stadien. Für die epimorphen Tiere allein lauten die Werte 0,15 /L in Fichtenstreu, 0,8 /L in Buchenlaub.) Diese Beobachtung steht im Einklang mit den Untersuchungsergebnissen zur Vertikalverteilung der Chilopoden und bestätigt die Vermutung, dass die Habitatstruktur für die räumliche Sonderung von L. mutabilis und L. (M.) curtipes eine Rolle spielt.
Experimentelle Eingriffe in die Waldboden-Lebensgemeinschaft
(Alle Ergebnisse sind nur als Arbeitshypothesen aufzufassen!)
12.3.1. Künstliche Populationen in 20 x 20 cm Plastikschachteln
(1) In gemischten Lithobius-Populationen überhöhter Dichte stellt sich eine ungefähr Freilandverhältnissen entsprechende Dichte und ein gleichbleibendes Verhältnis zwischen Beutezahl und Lithobiidenzahl ein.
(2) Lithobius mutabilis hat in den künstlichen Populationen die höchste Verlustquote und scheint der interspezifischen Konkurrenz weniger gewachsen zu sein als L. curtipes.
12.3.2. Lithobiiden unter Bodeneklektoren
(1) Unter Bodeneklektoren ist die Dichte der Mesofauna und der Lithobiiden nach einem Jahr verringert.
(2) Unter Bodeneklektoren ist nach einem Jahr die Dominanz von Lithobius mutabilis verringert.
(3) Unter Bodeneklektoren wird Lithobius curtipes von L. crassipes verdrängt. Dies ist ein Ausdruck größerer Trockenheit des Bodens.
12.3.3. Eingegrenzte Streuflächen
(1) Ein 7 - 8 cm in den Boden eingegrabener PVC-Ring bewirkt keine vollständige Isolation der eingeschlossenen Chilopoden.
(2) Eingegrenzte Streuflächen zeigen nach 2 bzw. 3 Jahren außerordentlich verschiedene Chilopodenabundanzen und -diversitäten. Dabei verändert sich die Populationsdichte von Lithobius mutabilis stärker als die von L. curtipes.
(3) Eine Vermehrung der unteren Streuschicht führt zu einer Diversitätssteigerung der Chilopoden, vor allem durch Steigerung der Evenness.
(4) Fichtenwald-Bodenstreu wird im Buchenwald so besiedelt, dass die für den Fichtenbestand typischen Abundanz- und Dominanzverhältnisse erhalten bleiben.
(5) Lithobius mutabilis bevorzugt Buchen-Fallaub gegenüber Fichtenstreu, bei Lithobius curtipes ist es umgekehrt.
Hinweis: Tabellen und Abbildungen können mit [Mausklick rechts -> Link in neuem Fenster öffnen] so aufgerufen werden, dass sie parallel zum Text geöffnet bleiben
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}