Das vorangegangene Kapitel zeigte, dass die Chilopodenfauna in verschiedenen Wäldern eine wiederkehrende taxonomische Grundstruktur aufweist, andererseits aber auch von Gebiet zu Gebiet deutliche Unterschiede in der Diversität und der Abundanzverteilung der Arten feststellbar sind. In diesem Kapitel wird die ökologische Sonderung verschiedener Hundertfüßlerarten in einem Gebiet näher analysiert. Als wesentliche Nischendimensionen werden die Variablen räumliche Verteilung, Nahrungsspektrum und zeitliche Verteilung betrachtet (SCHOENER 1974).
In dem Untersuchungsgebiet "Fabrikschleichach-Wald" (FW), einem ca. 140 Jahre alten Dentario-Fagetum auf Coburger Sandstein (470 müNN) wurden in den standardisierten Proben der Jahre 1979 bis 1983 insgesamt 2443 Chilopoden gefangen.
Sie repräsentieren elf Arten:
Lithobiomorpha
Lithobius mutabilis L. KOCH 1862: Das Verbreitungsgebiet der Art reicht von Nordfrankreich im Westen bis zu den Karpaten im Nordosten (VERHOEFF 1937). Eine Unterart von L. mutabilis findet sich auch in Griechenland (VERHOEFF 1925). In den Niederlanden und im Norden fehlt L. mutabilis. Die nördlichsten Funde in Deutschland sind das Niederbergische Land bei Wuppertal (THIELE 1956), der Solling (ALBERT 1982) und die Umgebung von Berlin (VERHOEFF 1925). L. mutabilis wird von VERHOEFF (1925) als Laubstreutier angesprochen, das nur ausnahmsweise außerhalb von Wäldern - vorzugsweise Buchenwäldern (THIELE 1956, LOKSA 1968) - zu finden ist. ALBERT (1982) und BROCKSIEPER (1972) fanden ihn jedoch auch auf Waldwiesen, in Kahlschlägen und Fichtenforsten.
L. muticus C.L. KOCH 1847: Das Verbreitungsgebiet dieser Art geht nach Norden etwas weiter als das von L. mutabilis. Es umfaßt auch noch die Niederlande (JEEKEL 1977), England EASON (1967) und Norddeutschland (Kiel) (TISCHLER 1980). Es handelt sich ebenfalls um ein Laubstreutier, anscheinend mit Vorliebe für Eichenwälder (THIELE 1956, BROCKSIEPER 1972). ALBERT (1982) fand L. muticus auch auf einer Waldwiese im Solling, und LOKSA (1966) stellte ihn als den dominanten Lithobiiden der xerothermen Flaumeichenwälder Ungarns fest.
L. macilentus L. KOCH 1862 (= L. aulacopus LATZEL 1880): Es handelt sich um eine Art mit Verbreitungsschwerpunkt in Nordwesteuropa, deren Vorkommen bis weit nach Skandinavien hinein reicht (MEIDELL 1979). Nördlich von Holland werden nur noch Weibchen dieser Art gefunden, so dass sie sich dort wahrscheinlich parthenogenetisch fortpflanzt (EASON 1964). BROCKSIEPER (1972) stellte fest, dass diese Art im Siebengebirge ihre höchsten Dichten in Waldbiotopen mittlerer Feuchte mit steinigem Boden und einem hohen Anteil an morschen Holz in der Streuschicht erreicht.
L. dentatus C.L. KOCH 1844: Diese Art hat wahrscheinlich ihren Verbreitungsschwerpunkt im südöstlichen Europa und findet bei Hamburg die Nordwestgrenze ihres Areals. Ihr Verbreitungsgebiet umfasst auch Nord- und Ostfrankreich, die Schweiz, die Tschechoslowakei, Ungarn und Jugoslawien (JEIKEL 1963). L. dentatus kommt regelmäßig in der Laubstreu von Wäldern vor (VERHOEFF 1925, BROCKSIEPER 1972), scheint aber nirgendwo häufig zu sein.
L. lusitanus valesiacus VERHOEFF 1935: Die (Unter-)art war bisher nur aus der Schweiz bekannt. (Die Nominatforn ist aus Portugal beschrieben.) Meine Funde aus dem Steigerwald stellen den ersten Nachweis für Deutschland dar (EASON 1982). EASON (pers. Mitt.) vermutet, dass L. lusitanus valesiacus in Mitteleuropa weiter verbreitet ist, bisher aber übersehen wurde.
L. pelidnus HAASE 1880: Die Art ist aus allen Ländern Nordwesteuropas bekannt (Skandinavien bis Österreich ohne Pyrenäen und Mittelmeerraum) (EASON 1987), scheint aber in ihrer Verbreitung weit nach Osten bis zum Kaukasus zu gehen (JEEKEL 1963). Es scheint sich um eine Waldart zu handeln, die in Mitteleuropa nur selten in hoher Abundanz vorkommt (BRÖLEMANN 1930, JEIKEL 1963).
Lithobius (Monotarsobius) curtipes C.L. KOCH 1847: Dies ist der am weitesten nach Norden vordringende Lithobiide überhaupt PALMEN 1948). Auch in Nordschweden finden sich Männchen und Weibchen von L. curtipes in gleicher Häufigkeit (TOBIAS 1975). Im Süden ist die Art aus Rumänien (MATIC 1966) und der Schweiz bekannt (EASON 1982). Sie scheint in Deutschland besonders feuchte oder gar moorige Wälder zu bevorzugen (VERHOEFF 1925).
L. (M.) crassipes L. KOCH 1862: Eine von Skandinavien bis Nordafrika, von England, Frankreich und Spanien bis nach Rußland, in den Kaukasus und nach Syrien verbreitete Art JEEREL 1963). L. crassipes kommt in der Laubstreu von Wäldern vor, wo JEEKEL (1977) ihn in den Niederlanden als das mehr trockenheitsliebende ökologische Gegenstück zu L. curtipes bezeichnet. Er findet sich auch häufig in morschem Holz (FAGER 1968, LLOYD 41963). In Kulturbiotopen lebt er unter Steinen und Brettern (WÜRMLI 1972). Im Steigerwald konnte ich die Art zusammen mit Lamyctes fulvicornis aus Wiesen-Bodenproben bestimmen.
L. (M.) aeruginosus L. KOCH 1862: Eine mitteleuropäische Art mit anscheinend kleinem Verbreitungsgebiet. Sie ist vor allem aus Deutschland (ohne Norddeutschland), Österreich, der Schweiz, Ungarn und Jugoslawien bekannt (EASON 1982, MATIC 1966). Im Siebengebirge ist L. aeruginosus eine der häufigsten Arten in Laubwäldern und offenem Gelände (BROCKSIEPER 1972). In den ungarischen Flaumeichenwäldern ist er der typische Vertreter der Lithobius-Untergattung Monotarsobius (LOKSA 1966).
Geophilomorpha
Strigamia acuminata (LEACH) (= Scolioplanes acuminatus LEACH): Die Art ist über ganz Europa von Skandinavien bis zum Kaukasus verbreitet (EASON 1964). S. acuminata bevorzugt die Laubdecke der Wälder und findet sich dort vor allen zwischen Blättern, in verrottendem Holz und unter Rinde (VERHOEFF 1925).
Geophilus spec. Eine unbestimmte Art (möglicherweise G. insculptus), die nur selten gefunden wurde.
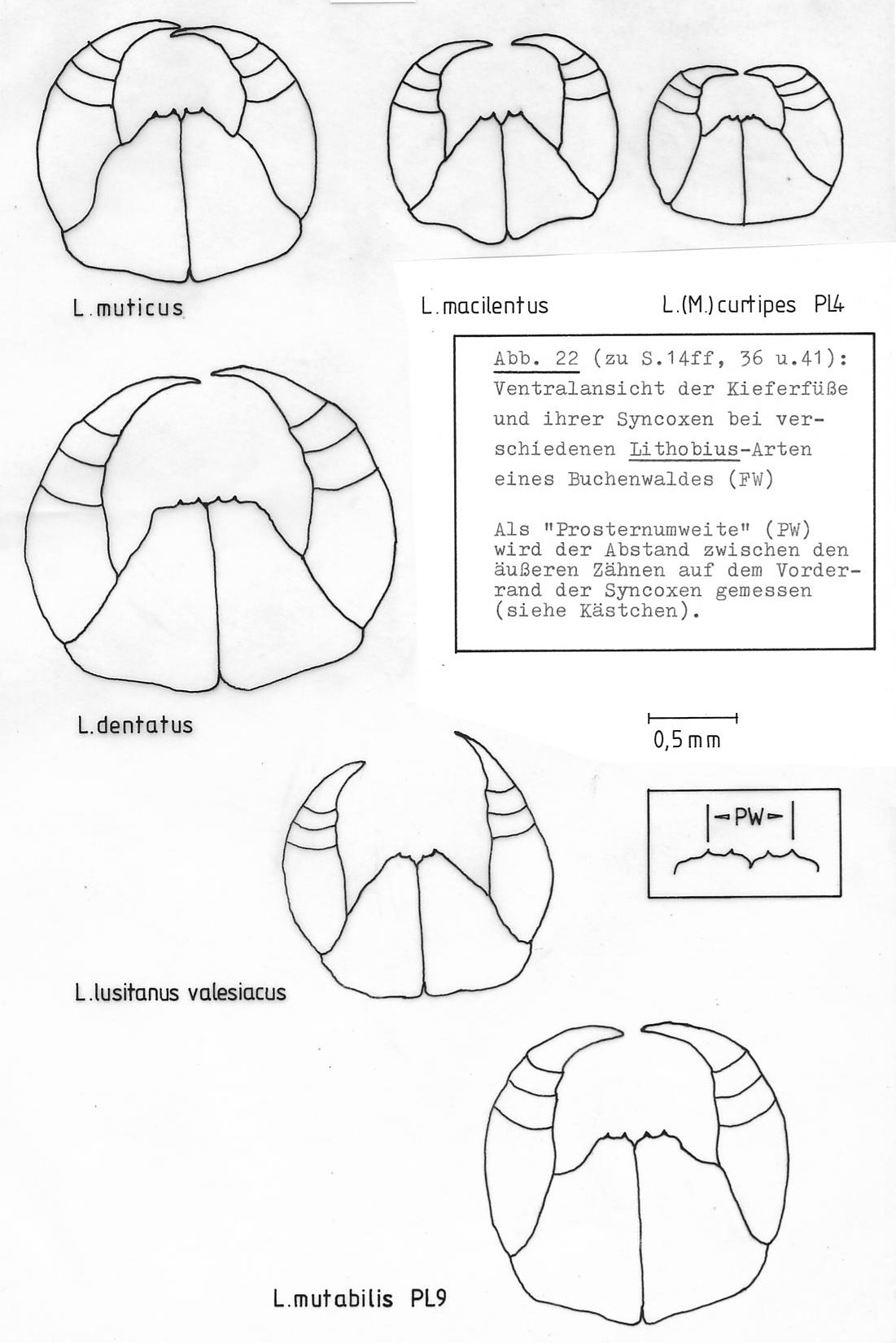
In Abb. 22 sind die Kieferfüße und ihre Syncoxen von adulten Tieren der Arten L. mutabilis, L. muticus, L.macilentus, L. dentatus, L. lusitanus valesiacus und L. curtipes abgebildet.
An 58 Quadratproben wurde für das Jahr 1979 geprüft, ob ein Zusammenhang zwischen der Lithobiiden-Artenzahl einer Probe und der Größe verschiedener am Probenort gemessener Habitatparameter (s. Tab.2) besteht. Das Ergebnis ist für 5 Parameter in Tab. 11 zusammengefasst,
Die Resultate dieses Vergleichs lassen natürlich nur Vermutungen über mögliche Zusammenhänge zu. Sie dienten aber zur Formulierung von Kriterien zur Standardisierung der Streuproben:
- Der nächste Stubben muss mindestens 2,50 m entfernt sein.
- Der nächste Stamm muss mindestens 2,50 m entfernt sein.
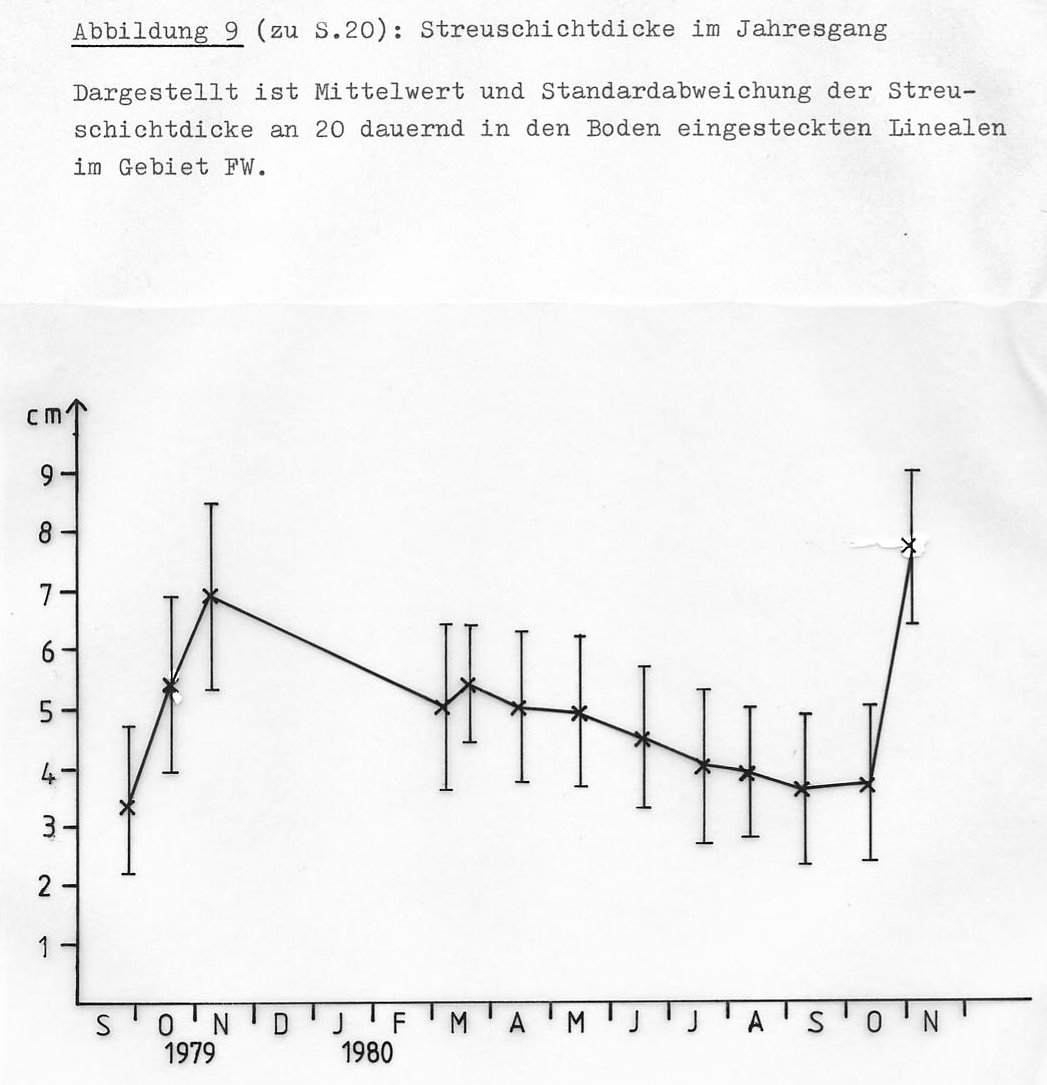
- Die Schichtdicke sollte zwischen 4,5 und 5,5 cm liegen. (Die Schichtdicke ändert sich im Jahresgang. Abb. 9 zeigt, dass die größten Dickenunterschiede im Spätherbst und Winter auftreten. Der Standardwert ist auf den Monat Juni bezogen und wurde für jeden Monat entsprechend der gemessenen Schichtdickenveränderung korrigiert.)
In allen Streuproben wurde die obere hauptsächlich unzersetzte Laubschicht (L-Horizont) getrennt von der unteren stärker zersetzten F-Schicht gesammelt und extrahiert. Zusammen mit der F-Schicht wurde auch alles lockere Bodenmaterial der Humusschicht aufgelesen. Streuproben dieser Art wurden monatlich von Februar bis Dezember 1980 genommen. Im September 1981 wurde die Laubstreu an der Basis von 4 Buchen und 4 Eichen extrahiert, im Oktober 1981 wurden 4 morsche Baumstümpfe ausgegraben und ebenfalls im Kempson-Apparat ausgelesen.
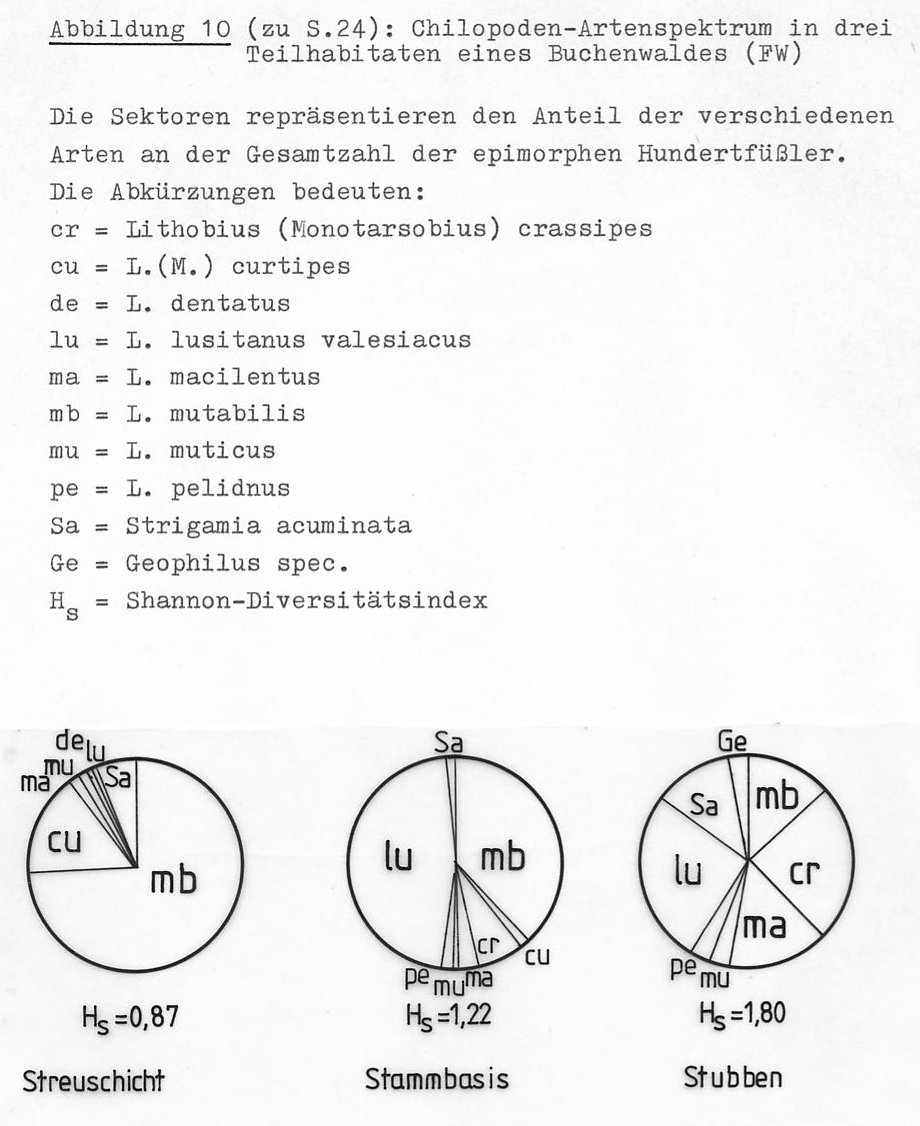
Abbildung 10 zeigt, dass in bezug auf die Chilopodenfauna große Unterschiede zwischen der reinen Laubstreu einerseits, Stubben und Stammbasen andererseits bestehen.
Der in der Streu nur sehr selten gefangene Lithobius lusitanus valesiacus scheint sein eigentliches Vorkommen im Stammbereich zu haben. Die beiden Arten Lithobius macilentus und L. (Monotarsobius) crassipes zeigen dagegen den größten Häufigkeitsanteil in den morschen Baumstümpfen, wo auch die Geophilomorphen mit zwei Arten und einem größeren Individuenanteil gefunden werden.
Lithobius pelidnus konnte in den Streuproben nicht gefunden werden, taucht aber im Stammbereich und in den Baumstubben auf. Die in der Streuschicht dominierende Art Lithobius mutabilis ist auch am Fuß der Baumstämmne und in den Stubbenproben nicht selten.
Einzelfänge von L. lusitanus valesiacus in den Fangschalen von Boden-Photoeklektoren und von L. pelidnus am Leimring um einen Buchenstamm sprechen dafür, dass diese Arten an Baumstämmen auf Nahrungssuche gehen.
In der freien Laubstreu wurden acht Chilopodenarten gefangen (Abb. 10 a). Nach Abzug der Arten, die schwerpunktmäßig in anderen Teilhabitaten des Waldes vorkommen (siehe oben), bleiben noch vier bis fünf Lithobiomorpha- und eine Geophilomorpha-Art, die bevorzugt oder ausschließlich in der Bodenstreu leben.
Es sind dies:
Lithobius mutabilis
L. dentatus
Lithobius (Monotarsobius) curtipes
Strigamia acuminata
Lithobius macilentus hat zwar seinen Verbreitungsschwerpunkt in den morschen Baumstümpfen, er ist aber in der Laubstreu so häufig - auch mit juvenilen Stadien -, dass sein Vorkommen dort wahrscheinlich nicht nur zufällig ist.
5.4.1.1. Häufigkeiten in oberer und unterer Streuschicht
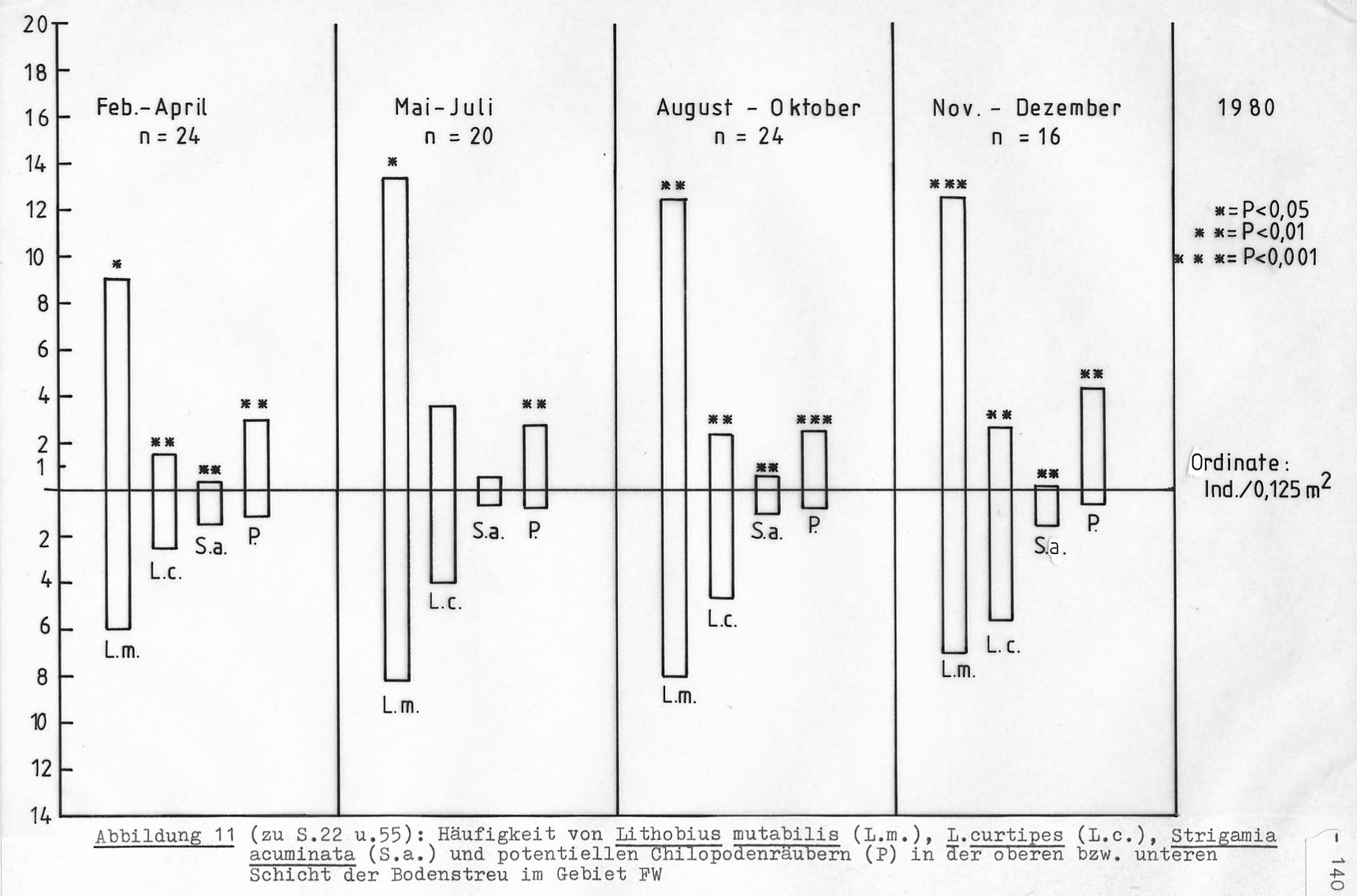
Die Fangzahlen erlauben nur für die häufigeren Arten eine Aussage zur Vertikalverteilung. Aus der Tabelle 12 ist zu erkennen, dass die dominierenden Arten deutlich unterschiedliche Schichten des Waldbodens bevorzugen.
Ein Blick auf die Abbildung 11 zeigt allerdings, dass sich daraus noch nicht ohne weiteres eine räumliche Trennung der verschiedenen Arten ergibt. Auch im unteren Schichtbereich ist Lithobius mutabilis der häufigste Chilopode. Diese Feststellung gilt auch, wenn nur die adulten Tiere betrachtet werden: In der F-Schicht haben adulte L. mutabilis eine Häufigkeit von 10 Tieren/m², adulte L. curtipes eine von 5 Tieren/m².
Im Mittel enthielt eine Quadratprobe der oberen Schicht 2,39+ 0,81 Arten, eine der unteren Schicht 2,93+ 0,88 Arten. Die Artendichte der Chilopoden ist also im unteren Streubereich deutlich höher als im oberen. (Signifikanzprüfung: t-Test; t = 4,14, P< 0,09)
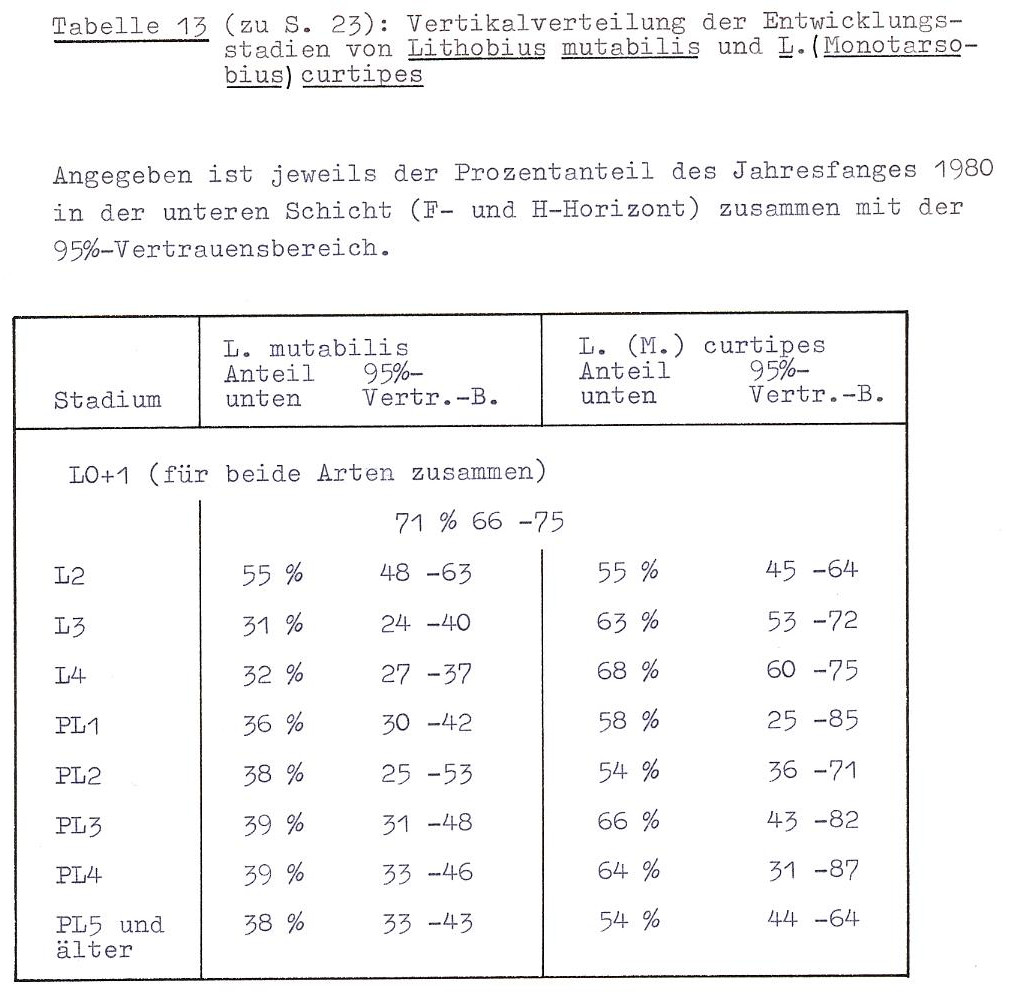
Betrachtet man die vertikale Häufigkeitsverteilung der Jugendstadien von L. mutabilis und L. curtipes (Tab. 13) so fällt auf, dass die L2-Stadien beider Arten gleichmäßig über den oberen und unteren Streubereich verteilt sind, während alle folgenden Entwicklungsstufen die gleichen Präferenzen wie die Adulten zeigen.
Bei der Durchsicht von 5 Streuflächen (20 x 20 cm²), die mit flüssigem Stickstoff vereist worden waren, bestätigten sich diese Ergebnisse: L. mutabilis fand sich in larvalen und postlarvalen Stadien zwischen zwei aufeinanderliegenden Blättern, neben kleinen Detritusanhäufungen zwischen den Blättern der Laubstreu und in der F-Schicht. Ein Weibchen saß in der oberen Humusschicht im horizontalen Grabgang eines Regenwurmes.
Von L. (M.) curtipes wurden insgesamt 6 Tiere in der unteren F-Schicht und im Übergang zwischen L- und F-Schicht beobachtet. L. curtipes saß mehr (4 Tiere) in schmalen Ritzen und Zwischenräumen, während die Aufenthaltsorte von L. mutabilis zu den Seiten hin in der Regel geräumiger waren.
In den vereisten Proben fand sich auch eine Strigamia acuminata in der oberen L-Schicht, die gerade einen Regenwurm gefangen hatte.
Vergleicht man die Ergebnisse von GEOFFROY (1979) und BOBERTS (1957) (2 Arbeiten mit vergleichbarer Erfassungsmethode) mit der im Gebiet FW festgestellten Vertikalverteilung, so fällt vor allem das fast vollständige Fehlen von Chilopoden in der L-Schicht der in Frankreich und England untersuchten Wälder auf. Der Unterschied ist zum Teil dadurch zu erklären, dass die von mir gesammelten Proben der oberen Schicht häufig auch etwas zersetztes Material enthielten. Die mit flüssigem Stickstoff vereisten Proben zeigen jedoch eindeutig, dass im Gebiet FW auch die reine L-Schicht von Chilopoden besiedelt ist. Entsprechend dem von mir festgestellten Verteilungsmuster im Gebiet FW fand ROBERTS eine Monotarsobius-Art (L. (M.) duboscqui = L. (M.) microps) in den tieferen, die Lithobius-Art (L. yariegatus) in den oberen Boden-Schichten.
In dem von ROBERTS untersuchten englischen Buchen-Eichenwald sind die Unterschiede in der Schichtenpräferenz wesentlich stärker ausgeprägt als bei den Chilopoden-Arten im Steigerwald.
5.4.1.2. Lässt sich die unterschiedliche Vertikalverteilung der Chilopodenarten durch ihr Verhalten gegenüber verschiedenen Luftfeuchten erklären?
5.4.1.2.1. Feuchtepräferenz
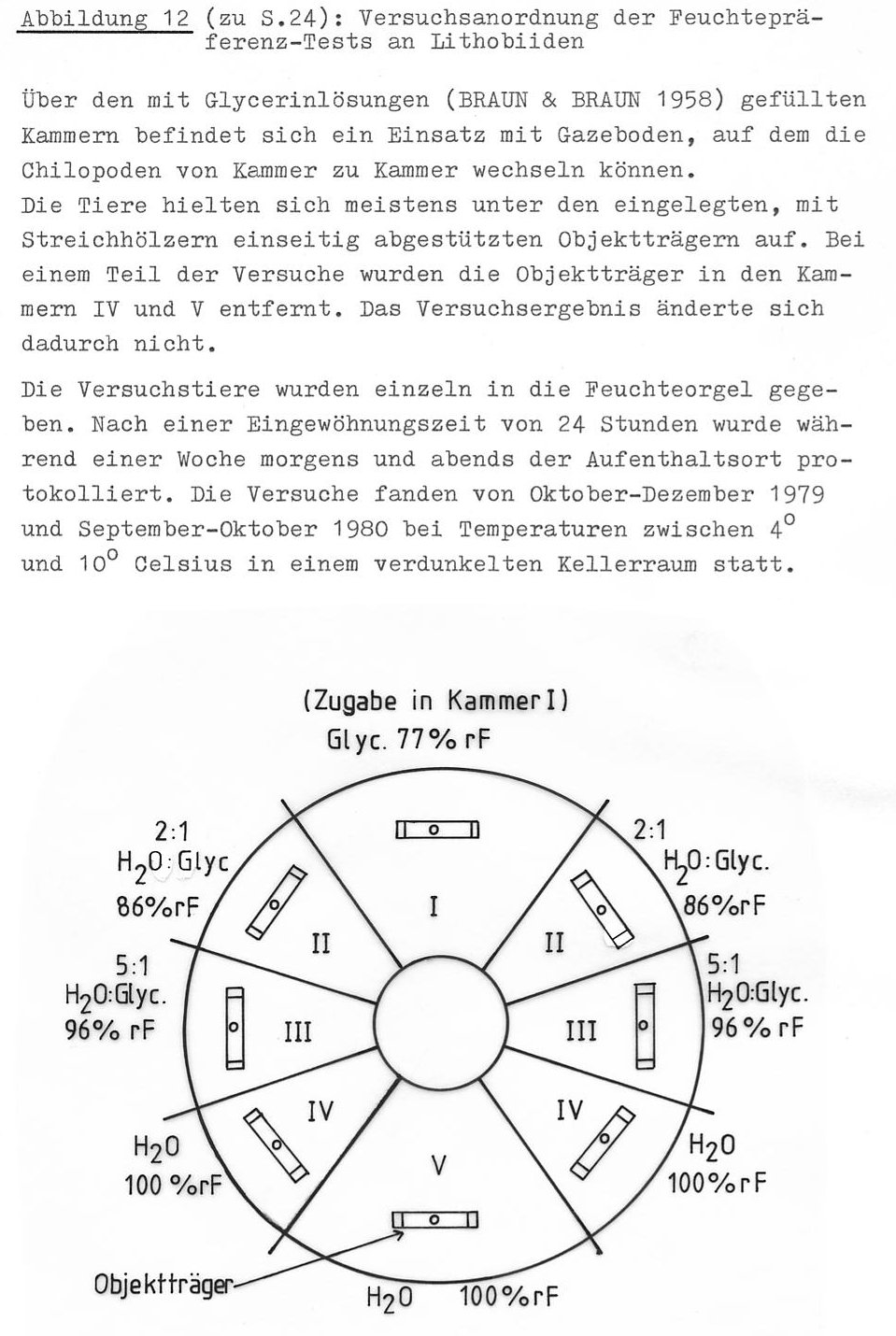
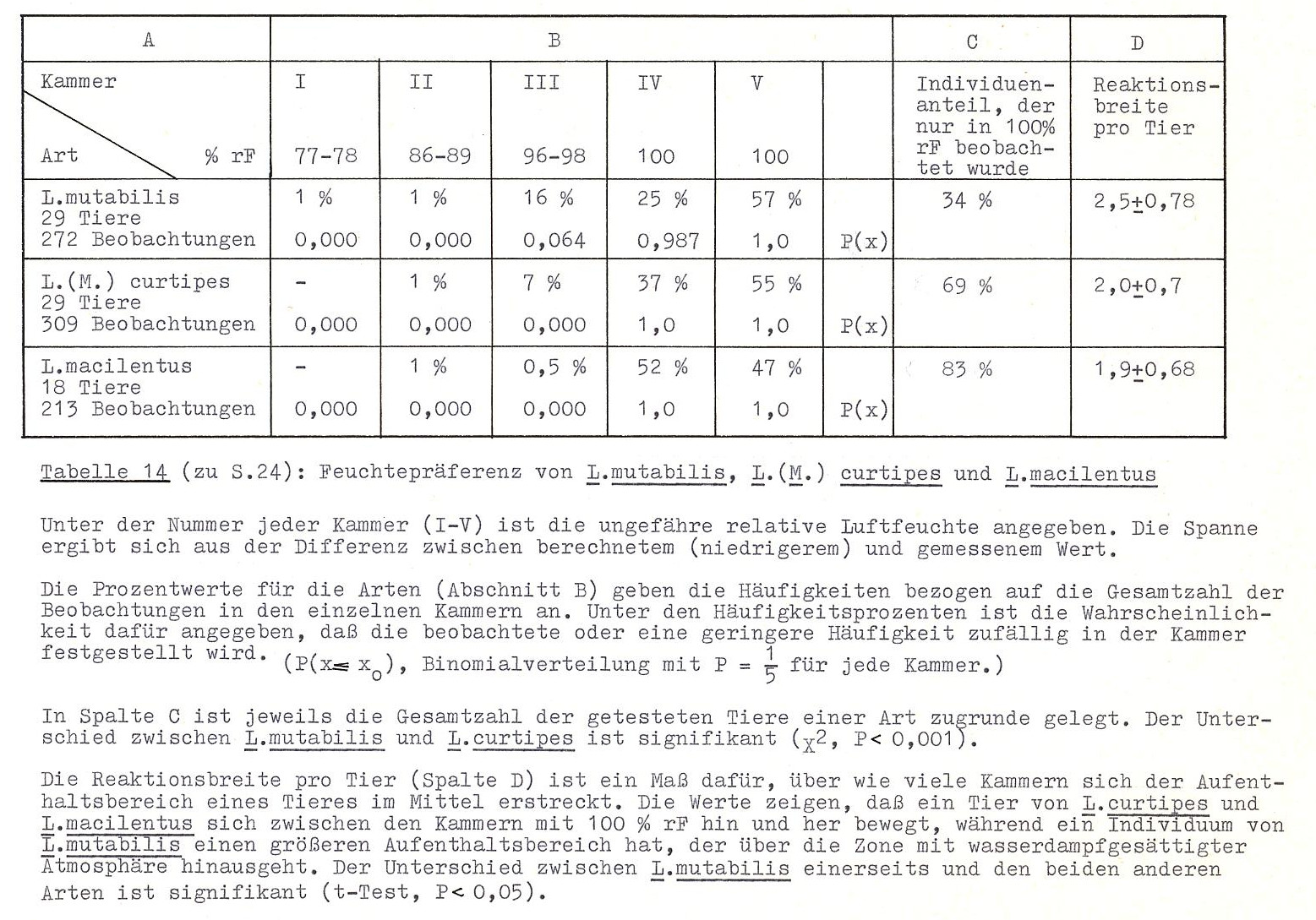
Für die Arten Lithobius mutabilis, L. (M.) curtipes und L. macilentus wurde die Luftfeuchtepräferenz in Feuchteorgeln mit Glycerin-Wasser-Gemisch verschiedener Konzentrationen geprüft (Abb. 12). Die Ergebnisse sind in Tab. 14 dargestellt. Es zeigt sich, dass alle Arten wasserdampfgesättigte Atmosphäre aufsuchen. Dabei meiden L. curtipes und L. macilentus sogar schon Feuchten von 96 - 98 % rF als Aufenthaltsort. (Die Wahrscheinlichkeit, dass dieses Ergebnis auf Zufall beruht, ist kleiner als 10-9.) Lithobius mutabilis war im Feuchtebereich von 96 - 98 % rf signifikant häufiger vertreten als die beiden anderen Arten, muss aber dennoch eindeutig als 100 % rF bevorzugend angesprochen werden.
In Anbetracht der Tatsache, dass die Chilopoden über keinen oder nur einen sehr geringen Verdunstungsschutz verfügen (BLOWER 1951, CURRY 41974) sind diese Ergebnisse nicht verwunderlich und entsprechen auch den Befunden von ALBERT (im Druck a) und AUERBACH (1949, 1951).
5.4,1.2.2. Trockentoleranz
Größere Unterschiede zwischen den Arten zeigen sich in der Toleranz gegenüber nicht-wasserdampfgesättigter Atmosphäre. Das Ergebnis der Versuche zur Trockentoleranz ist in Tabelle 15 zusanmengefasst.
Die Unterschiede in der Trockentoleranz entsprechen weitgehend der Vertikalverteilung der verschiedenen Arten: L. mutabilis weist die größte Trockentoleranz auf, während die auf die tieferen Streuschichten beschränkten L. curtipes und Strigamia acuminata in trockener Luft wesentlich kürzer überleben. Generell sind größere Tiere in der Lage, längere Trockenzeiten zu überstehen als kleinere Tiere, Dies hat geometrische Ursachen im Oberflächen-Volumen-Verhältnis. Jedoch sind auch die größengleichen Individuen von L. mutabilis und L. curtipes in ihrer Trockentoleranz verschieden. Das zeigt, dass L. mutabilis physiologisch besser an das Überstehen von (kurzen) Zeiten in einer Atmosphäre mit Feuchtedefizit angepasst ist.
Ich machte die Beobachtung, dass durch Austrocknung bewegungsunfähige Lithobiiden in der Lage sind, nach Wiederbefeuchtung Wasser aufzunehmen und in voller Vitalität weiterzuleben. Diese Revitalisierung ist aber nur möglich, wenn die Austrocknung nicht zu weit fortgeschritten ist. BLOWER (1955) erwähnt diesen Vorgang ebenfalls. Nach seinen Angaben tritt die Trockenstarre bei ca. 20 % Gewichtsverlust ein. Bei Wasseraufnahme nur durch die dorsale Cuticula ist das ursprüngliche Gewicht in 6 Stunden wieder erreicht.
ROBERTS (1957) führte vergleichbare Versuche zur Trockentoleranz an Lithobiiden durch (L. variegatus, L. lapidicola, L. (M.) duboscqui = L. (M.) microps (EASON 1982)). Die Ergebnisse entsprechen größenordnungsmäßig den Werten, die ich bei den von mir untersuchten Arten bestimmte. ROBERTS fand jedoch wesentlich größere Unterschiede in der Trockentoleranz zwischen den beiden Hauptarten seines Gebietes: L. (M.) microps mit Überlebenszeit 3,3 Stunden bei 66 % rF und 19 °C gegenüber 15,2 Stunden für größengleiche L. variegatus. Dieses Ergebnis passt gut zu der oben erwähnten stärkeren vertikalen Sonderung der Lithobiiden in dem von ROBERTS untersuchten Wald.
5.4.2.1. Verteilungsmuster der Arten
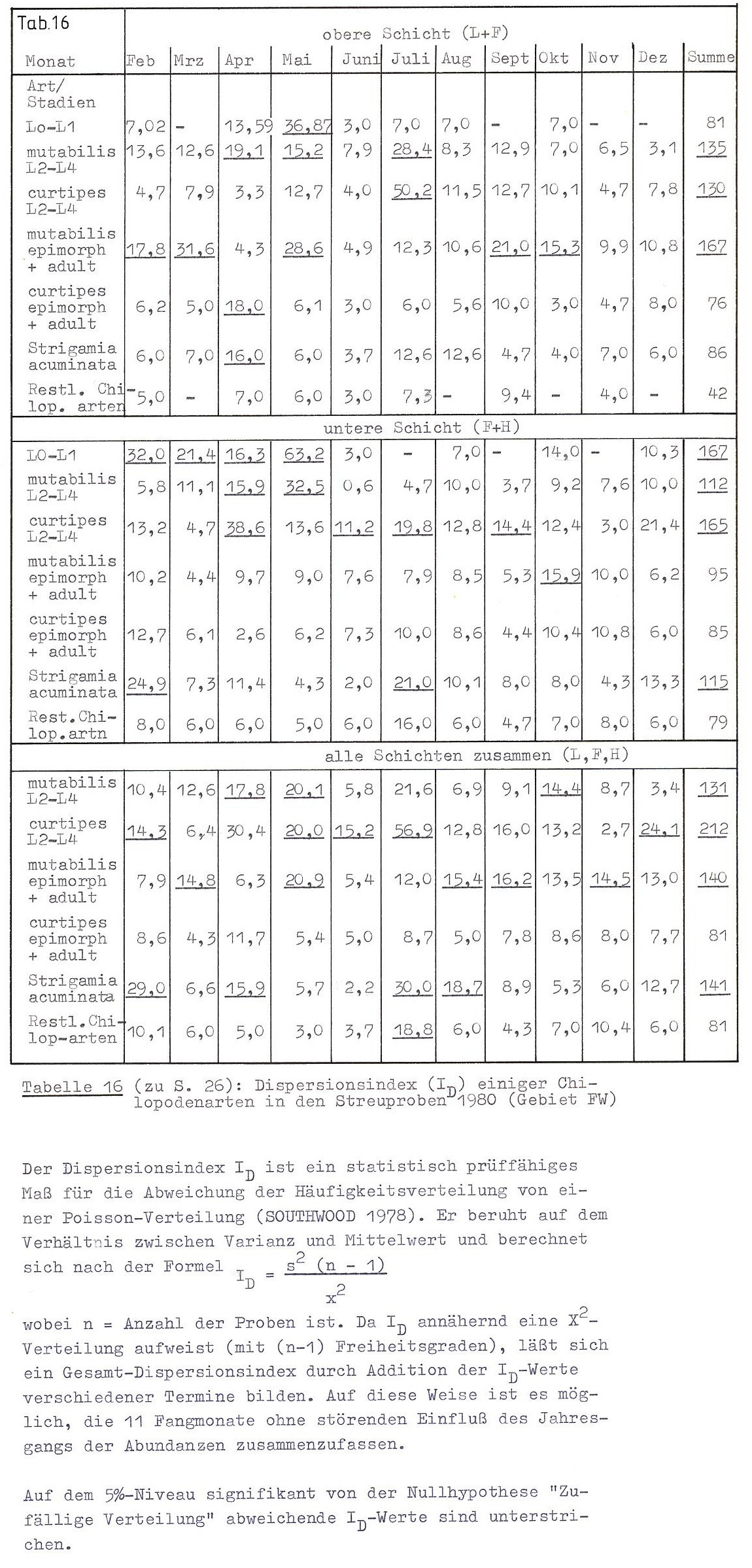
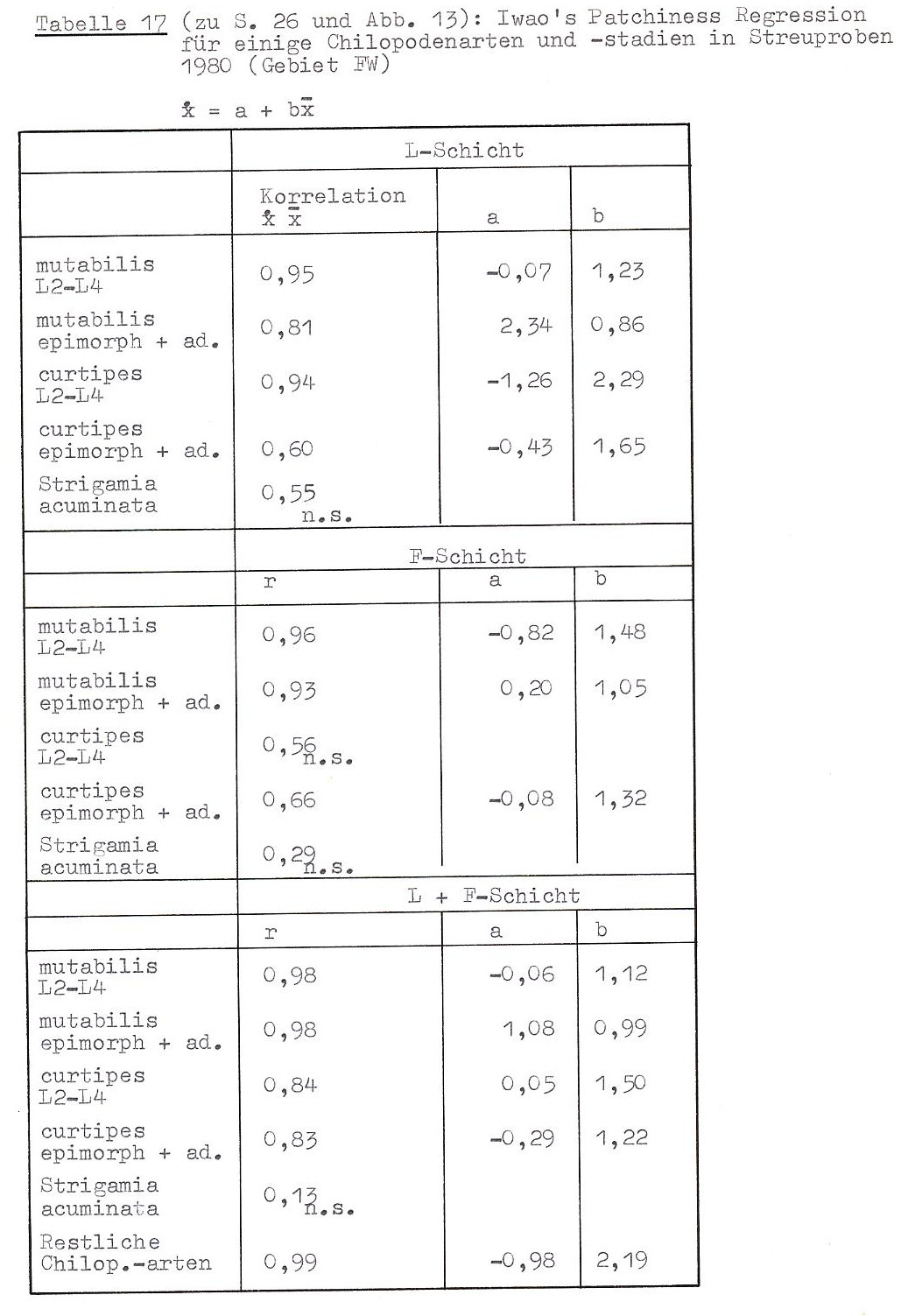
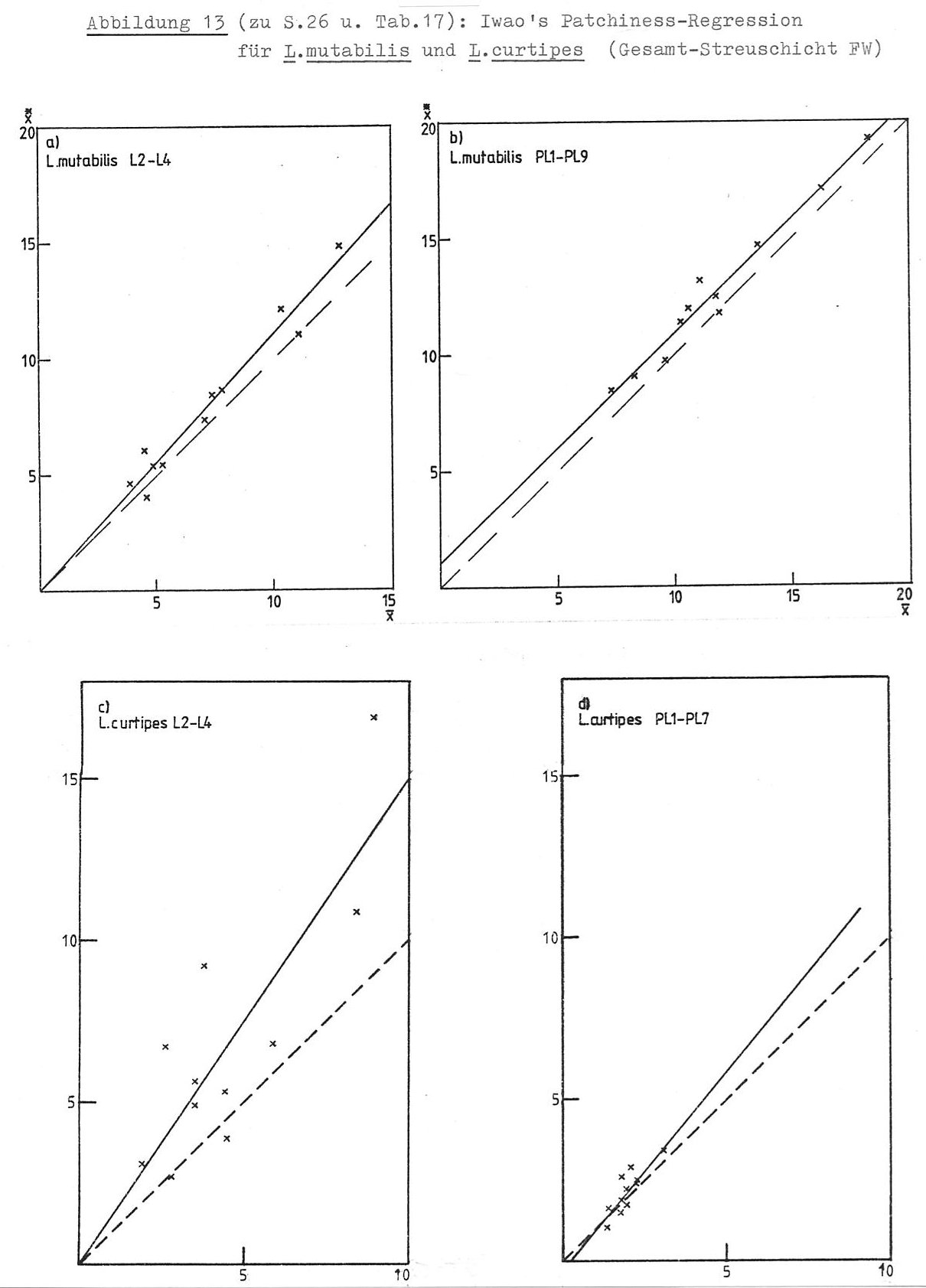
Um zu prüfen, ob die Chilopoden in dem von mir untersuchten Gebiet zufällig verteilt sind, berechnete ich die in Tabelle 16 dokumentierten ID-Werte. (Zu Formel und Eigenschaft des Dispersionsindex ID siehe die Legende zu Tab. 16.) Es zeigen sich folgende Ergebnisse:
Die untersuchten Chilopoden sind entweder zufällig oder kumulativ verteilt. Eine kumulative Verteilung haben die anamorphen Stadien von L. (M.) curtipes, alle Stadien von L. mutabilis sowie Strigamia acuminata. L. (M.) curtipes ist in den epimorphen und adulten Stadien zufällig verteilt. Über die restlichen Chilopodenarten ist keine Aussage möglich, da bei ihnen die Zufallsverteilung sich (auch?) aus der großen Zahl "leerer" Proben ergibt.
Bei zwei Arten zeigen sich Unterschiede im Verteilungsmuster zwischen oberer und unterer Bodenschicht: Epimorphe und adulte L. mutabilis sind nur in der oberen Schicht aggregiert, im unteren Bodenbereich jedoch zufällig verteilt. Umgekehrt ist die kumulative Verteilung von Strigamia acuminata nur in der F- und H-Schicht festzustellen.
Die Daten legen die Interpretation nahe, dass die anamorphen Stadien von L. mutabilis und L. (M.) eurtipes ohne direktes Aggregationsverhalten aufgrund externer Faktoren (Habitatmosaik?) kumuliert sind (a ≈ 0, b >1), während die adulten und epimorphen L. mutabilis Individuengruppen bilden, die selbst zufällig verteilt sind (a > 0, b ≈ 1). REISE & WEIDEMANN (1975) kommen durch Auszählen eines Rasters zusammenhängender Isolierquadrate (156 cm²) im Sauerhumus-Buchenwald des Solling ebenfalls zu dem Ergebnis, dass epimorphe und adulte L. (M.) curtipes bei allen Quadratgrößen zufällig verteilt sind, während epimorphe und adulte L. mutabilis Aggregationen bis zu einer Fläche von weniger als 300 cm² bilden, die selbst zufällige Verteilung aufweisen. Anamorphe Stadien der Lithobiiden sowie Geophilomorpha wurden von REISE & WELDEMANN nicht untersucht.
5.4.2.2. Assoziationen zwischen den Arten
Die sehr hohe Präsenz der beiden häufigsten Chilopodenarten in den 50 x 25 cm²-Proben (L. mutabilis 100 %, L. (M.) curtipes 96 %) macht es sinnlos, Assoziationen zwischen ihnen bzw. ihnen und den selteneren Arten zu berechnen. Die gegenseitige Unabhängigkeit des Vorkommens wurde deshalb nur für die Arten L. muticus, L. macilentus und Strigamia acuminata geprüft. Im Vierfeldertest (SOUTHWOOD 1978, MÜHLENBERG 1976) erlaubt der X² -Wert für keines der drei Artenpaare ein Zurückweisen der Nullhypothese: "Die Arten sind in ihrem Vorkommen unabhängig voneinander".
Die Prüfung auf Assoziation nach FAGER (1957) (Berücksichtigung nur der Proben, in denen mindestens eine der geprüften Arten vertreten ist) ergibt für das Artenpaar L. Macilentus/ L. muticus einen t-Wert von -3,17, dessen Betrag weit über der Simnifikanzschwelle (5%-Niveau) von t = 1,96 liegt. (Tab. 18) Ich halte es für berechtigt, dieses Ergebnis im Sinne einer negativen Assoziation zwischen den beiden Arten zu interpretieren. FAGER (1957) wendet gegen die Prüfung negativer Assoziationen mit seinem Verfahren ein, dass "Nicht-Finden" einer Art eine methodisch zu unsichere Sache sei, um daraus Schlüsse zu ziehen. Im vorliegenden Fall ist die Sicherheit aber nahezu hundertprozentig, dass eine nicht aus den 25 x 50 cm² Flächenproben extrahierte Art dort auch tatsächlich nicht vertreten war.
Um zwischen den dominanten Arten positive oder negative Beeinflussungen feststellen zu können, berechnete ich verschiedene Korrelationskoeffizienten für den Zusammenhang zwischen den Häufigkeiten der Arten in den Streuproben. (Produkt-Moment-Korrelation r, Spearman-Rangkorrelation rs, Kendall's tau). Ein Einfluss der Phänologie wurde durch getrennte Berechnung der Korrelationen für die 4 Quartale des Jahres (Feb.-April, Mai-Juli, Aug.-Okt., Nov.-Dez.) weitgehend ausgeschlossen. Es fanden sich keine eindeutigen Korrelationen.
Käumlichs Verteilung der Chilopoden in einem Buchenaltbestand.
A, Teilhabitate
(1) Die Teilhabitate Stubben und Stammbasen enthalten eine Chilopodenfauna, die sich deutlich von der der freien Laubstreu unterscheidet.
(2) Von den 11 im Bestand gefundenen Chilopodenarten können 2 den Stubben und 2 den Stammbasen als Hauptlebensraum zugeordnet werden.
B, Vertikalverteilung
(1) Die beiden dominanten Chilopodenarten Lithobius mutabilis und L. curtipes unterscheiden sich in ihrer Vertikalverteilung im Waldboden; der Unterschied zeigt sich bereits bei den anamorphen Jugendstadien.
(2) L. mutabilis ist trotz seiner Präferenz für die obere Streuschicht auch im unteren Streubereich die häufigste Art, so dass sich aus der verschiedenen Vertikalverteilung keine einfache räumliche Trennung der Arten ergibt.
(3) Die Chilopoden-Artendichte ist in der unteren Streuschicht höher als in der oberen.
(4) Die unterschiedliche Vertikalverteilung der Lithobius-Arten 1äßt sich nicht ohne weiteres durch verschiedene Feuchtepräferenzen erklären: Im Wahlversuch bevorzugen die Arten Lithobius mutabilis, L. (M.) curtipes und L. macilentus alle 100 % relative Luftfeuchte, die Amplitude der aufgesuchten Feuchtebereiche ist jedoch bei L. mutabilis größer als bei den beiden anderen Arten.
(5) Lithobius mutabilis kann trockene Luft länger ertragen als L. curtipes und Strigamia acuminata; dies gilt auch für größengleiche Individuen der drei Arten.
C, Horizontalverteilung
(1) Die in der freien Streuschicht dominanten Arten unterscheiden sich im horizontalen Verteilungsmuster: Lithobius mutabilis bildet Aggregationen, L. curtipes ist zufällig verteilt.
(2) Zwischen den beiden subdominanten Arten Lithobius muticus und L. macilentus besteht eine negative Assoziation. Die drei dominanten Arten (L. mutabilis, L. curtipes, Strigamia acuminata) scheinen in ihrem horizontalen Verteilungsmuster unabhängig voneinander zu sein.
Hinweis: Tabellen und Abbildungen können mit [Mausklick rechts -> Link in neuem Fenster öffnen] so aufgerufen werden, dass sie parallel zum Text geöffnet bleiben
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}